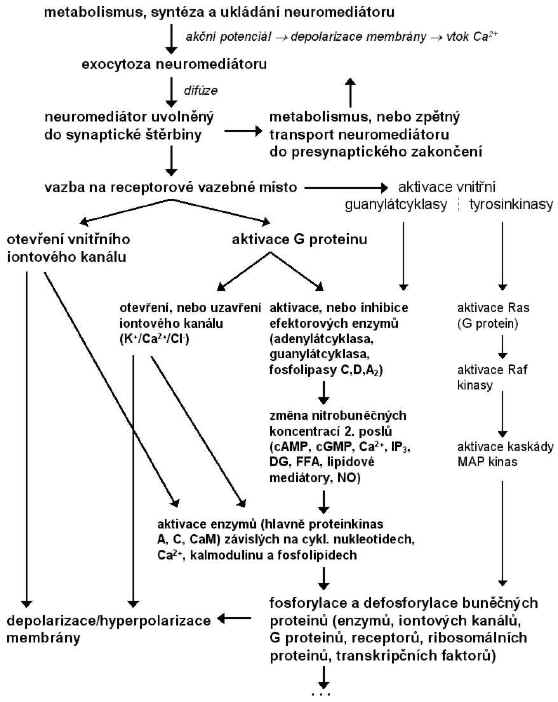
Neurotransmisí se obecně rozumí přenos nervového signálu podél neuronů i mezi nimi. Transdukce je převod nervového signálu z akčního potenciálu na signál jiné fyzikálně-chemické podstaty. Na Obr. 6 je zjednodušené schéma hlavních stupňů v převodu signálu přes chemickou synapsi. Receptory s enzymovou aktivitou, které jsou ve schématu rovněž uvedeny se na synaptickém přenosu nepodílejí přímo; jejich agonisty jsou růstové faktory, cytokiny a některé hormony. Přenos signálu je přirovnáván k 3-rozměrné síti, v níž podráždění v jednom bodě se přenese do všech dalších. Existují důkazy o tom, že např. regulace snížením počtu b-AR, jakožto obvykle pozorovaný účinek dlouhodobého podávání antidepresiv, může být zprostředkována křížovým propojením receptorů přes PKC.
Přenos signálu mezi povrchově lokalizovaným membránovým receptorem a příslušným efektorovým enzymem (např. AC nebo PLC) je uskutečňován prostřednictvím regulačního prvku (transduceru), kterým je některý z G proteinů. Odhaduje se, že kolem 80% primárních poslů (hormonů, neuromediátorů, neuromodulátorů, buněčných aktivačních faktorů) působí přes receptory spojené s G proteiny (36).
Strukturně lze G proteiny rozdělit na monomerní a trimerní. Malé G proteiny jsou zřejmě zapojeny v metabolismu fosfolipidů. Heterotrimerní G proteiny sestávají z podjednotek a, b, g, které mají řadu izotypů. a podjednotky (Ga) nesou vazebné místo pro guaninové nukleotidy (GDP, GTP) a mají vlastní GTPasovou aktivitu. Specifické působení G proteinů na efektorovou odezvu je obvykle zajištěno aktivovanou Ga (s navázaným GTP), ale uplatňovat se mohou i bg komplexy. Ga existují v rodinách Gas, Gaq a Gai, z nichž každá má další větve a ty mají jednotlivé členy (325, 194, 226). V přenosových systémech monoaminových receptorů se uplatňují především tyto skupiny G proteinů: 1) Gs stimulující AC a tedy i tvorbu cAMP; 2) Gi inhibující AC a Go působící na AC, fosfolipasy i iontové kanály; 3) Gq a G11 regulující aktivitu PLC a tedy hydrolýzu PIP2. Rovněž b a g podjednotky mají řadu podtypů. Samotné bg komplexy zřejmě aktivují PLC (44) a předpokládá se, že bg dimery uvolněné aktivací Gi izoforem mohou regulovat PLA2 a tedy ovlivňují metabolismus membránových fosfolipidů spojený s přenosem signálu. Na některých dějích, jako otevírání K+-kanálů, se zřejmě mohou podílet a podjednotky i bg dimery. Navíc byl identifikován membránový protein, který zřejmě reguluje aktivaci G proteinu tím, že stabilizuje komplex agonista-receptor-G protein. Tento připojený kofaktor může mít úlohu ve křížovém propojení receptorů a ve vylaďování signálu.
Reverzibilní fosforylace proteinů je pravděpodobně nejdůležitějším molekulárním mechanismem, kterým mimobuněčné signály realizují biologickou odezvu v cílových neuronech. Mezi neuromediátory, které mohou působit přes fosforylaci/defosforylaci proteinů patří např. acetylcholin, noradrenalin, dopamin, serotonin, adenosin, glutamát a další. Významná je hlavně fosforylace iontových kanálů, receptorů, enzymů zahrnutých do syntézy neuromediátorů, proteinů spojených se synaptickými váčky (synapsinů), cytoskeletálních proteinů a inhibitorů fosfatas. Fosforylace, tj. přenos zbytku kyseliny fosforečné z ATP na hydroxylové skupiny serinu, threoninu nebo tyrosinu v proteinech, je katalyzována proteinkinasami. Fosfatasy defosforylují fosfoproteiny a vracejí systém do základního stavu. Různé proteinkinasy jsou fyziologicky aktivovány 1) druhými posly (cAMP, cGMP, IP3, DG, Ca2+, kyselinou arachidonovou, lipidovými mediátory); 2) autofosforylací proteinkinas; 3) vazbou agonistů k receptorům spojeným s proteinkinasami (obvykle receptory s tyrosinkinasovou aktivitou, např. receptory pro růstové faktory, hormony a mitogeny); 4) bg podjednotkami G proteinů. Různé proteinkinasy přitom mohou ovlivňovat tentýž systém. Např. rychlá fosforylace receptorů spojených s G proteiny, která často doprovází jejich desenzibilizaci, může být způsobena dvěma typy kinas - proteinkinasami zavislými na druhých poslech a protenkinasami receptorů spojených s G proteiny (304, 164).
Obr. 6: Hlavní stupně přenosu synaptického přenosu signálu
Podporováno grantem GA UK č. 51/1996.