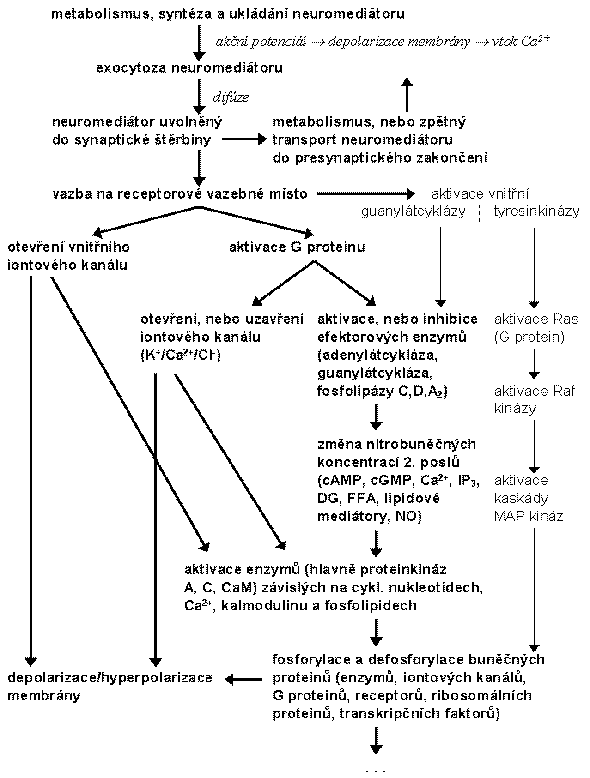
Hlavní stupně synaptického přenosu signálu
Dnes je přenos signálu přirovnáván k 3-dimenzionální síti, v níž podráždění v jednom bodě se přenese do všech dalších. Skutečný stav může vypadat takto: adenylátcykláza je současně aktivována i inhibována; dva různé receptory mohou působit opačně na Ca2+-kanál; může se potencovat uvolnění Ca2+ z nitrobuněčných rezerv a vtok zvnějšku atd.
Na obrázku je příklad křížového propojení systémů receptor-efektor mezi receptory aktivujícími obrat fosfoinositidů a receptory spojenými s adenylátcyklázou. Aktivace fosfolipázy C vede ke generaci IP3 (® mobilizace Ca2+) a DG a tedy k aktivaci proteinkinázy C (PKC). Existuje řada míst pro možnou interakci PKC a systémů regulujících produkci cAMP:
fosforylace b-adrenergního receptoru vede k jeho odpojení od Gs;
fosforylace adenylátcyklázy způsobuje zvýšení její aktivity;
fosforylace Gi umožňuje inaktivaci inhibičních účinků.
Výsledný účinek závisí na rozdílech mezi podtypy G proteinů a na relativní početnosti izoenzymů proteinkinázy C.
Dalším příkladem je vzájemné ovlivnění afinity receptorů G proteiny, umožněnou relativní zaměnitelností bg podjednotek. bg komplex z Gs je velmi podobný nebo identický jako u Gi ® uvolnění bg komplexu (např. aktivací D1 receptoru) může ovlivnit rovnováhu mezi disociovaným a nedisociovaným stavem u Gs proteinu a tím modulovat afinitu jiných receptorů (např. D2 receptoru, a2-adrenoceptoru).
Pozn.:
1) Existují důkazy o tom, že downregulace (= regulace snížením počtu) b-adrenergních receptorů (jakožto obvykle pozorovaný účinek dlouhodobého podávání antidepresiv) může být zprostředkován křížovým propojením přes proteinkinázu C.
Byl identifikován membránový protein, který reguluje receptorem zprostředkovanou aktivaci G proteinu tím, že stabilizuje komplex agonista-receptor-G protein. Tento připojený kofaktor může mít úlohu ve křížovém propojení receptorů a ve vylaďování signálu.
Reverzibilní fosforylace proteinů je pravděpodobně nejdůležitějším molekulárním mechanismem, kterým mimobuněčné signály realizují biologickou odezvu v cílových neuronech. Mezi neuromediátory, které mohou působit přes fosforylaci/defosforylaci proteinů patří např. acetylcholin, noradrenalin, dopamin, serotonin, adenosin, glutamát a další. Významná je hlavně fosforylace iontových kanálů, receptorů, enzymů zahrnutých do syntézy neuromediátorů, proteinů spojených se synaptickými váčky (synapsinů), cytoskeletálních proteinů a inhibitorů fosfatáz.
Základní pojmy:
Fyziologické mechanismy aktivace proteinkináz:
druhými posly (cAMP, cGMP, IP3, DG, Ca2+, kys. arachidonová, lipidové mediátory);
autofosforylací proteinkináz;
vazbou agonistů k receptorům spojeným s proteinkinázami (obvykle s tyrosinkinázovou aktivitou, např. receptory pro růstové faktory, hormony a mitogeny).
Hlavní
fyziologické funkce proteinkináz v mozku
|
typ proteinkinázy |
hlavní fyziol. funkce |
|
|
|
|
závislá na cAMP |
regulace iontového kanálu |
|
syntéza mediátoru |
|
|
uvolňování mediátoru |
|
|
desenzitizace receptoru |
|
|
intermediární metabolismus |
|
|
|
|
|
závislá na cGMP |
regulace iontového kanálu |
|
|
|
|
CaM-II (závislá na Ca2+ a kalmodulinu) |
syntéza mediátoru |
|
uvolňování mediátoru |
|
|
LTP |
|
|
organizace cytoskeletu |
|
|
|
|
|
proteinkináza C (závislá na Ca2+ a fosfolipidu) |
regulace iontového kanálu |
|
syntéza mediátoru |
|
|
uvolňování mediátoru |
|
|
desenzitizace receptoru |
|
|
LTP |
Hlavním způsobem zvýšení fosforylace je zvýšení aktivity proteinkináz, ale do regulačních mechanismů mohou být zapojeny i různé fosfatázy a změny substrátových proteinů. Řada proteinů může přitom být fosforylována více různými proteinkinázami. V CNS existují vyšší aktivity proteinkináz závislých na cAMP a na Ca2+ a větší počet možných substrátových proteinů, než je tomu v periferních tkáních.
A) Proteinkinázy závislé na cyklických nukleotidech
PK závislé na cAMP
PK závislé na cGMP
B) Proteinkinázy závislé na Ca2+/kalmodulinu
PK II závislá
na Ca2+/kalmodulinu
PK I závislá
na Ca2+/kalmodulinu
PK III závislá
na Ca2+/kalmodulinu
Proteinkinázy
v mozku
|
Proteinkinázy
regulované druhými posly |
|
|
Proteinkinázy závislé na cyklických nukleotidech |
PK závislé na cAMP (proteinkinázy A) |
|
PK závislé na cGMP |
|
|
Proteinkinázy závislé na Ca2+/kalmodulinu |
PK II závislá na Ca2+/kalmodulinu |
|
PK I závislá na Ca2+/kalmodulinu |
|
|
PK III závislá na Ca2+/kalmodulinu a další |
|
|
Proteinkinázy závislé na Ca2+ a fosfolipidu
(proteinkinázy C) |
|
|
Proteinkinázy
nezávislé na druhých poslech |
|
|
|
kaseinkináza I a II |
|
|
proteinkinázy neurofilament |
|
|
kináza myelinovéhobasického proteinu |
|
|
kináza pyruvát dehydrogenázy a další |
|
Tyrosin-specifické
proteinkinázy |
|
|
Protoonkogenní produkty |
|
|
Receptory růstových faktorů (s vnitřní tyrosinkinázovou aktivitou) |
|
(podle Walaas S.I. and Greengard P.: Pharmacol. Rev. 43 (3) 299-349, 1991)
C) Proteinkinázy závislé na Ca2+ a
fosfolipidu (PKC)
Mozek obsahuje řadu dalších proteinkináz, které nejsou závislé na druhých poslech a které fosforylují mozkové proteiny na serinových a threoninových zbytcích (kaseinkináza I a II, kináza 3 glykogen syntázy, proteinkinázy neurofilament, kináza basického myelinového proteinu, kináza pyruvát dehydrogenázy atd.) a dále tyrosin-specifické kinázy (třída protoonkogenních produktů a třída receptorů růstových faktorů s vnitřní tyrosinkinázovou aktivitou).
Izoformy PKC
|
typ |
podtyp |
aktivátory |
tkáňová exprese |
|
cPKC
(konvenční) |
a |
Ca2+,
DG, PS, |
univerzální |
|
|
FFA,
lysoPC |
|
|
|
bI |
FFA,
lysoPC |
některé
tkáně |
|
|
bII |
FFA,
lysoPC |
hodně
tkání |
|
|
g |
FFA,
lysoPC |
pouze
mozek |
|
|
nPKC
(nové) |
d |
DG,
PS |
univerzální |
|
e |
DG,
PS, FFA, PIP3 |
mozek
a jiné |
|
|
h
(L) |
DG,
PS, PIP3, |
kůže,
plíce, srdce |
|
|
|
cholesterolsulfát |
|
|
|
Q |
? |
sval,
T buňky aj. |
|
|
m |
? |
NRK
buňky |
|
|
aPKC
(atypické) |
z |
PS,
FFA, PIP3? |
univerzální |
|
l
(i) |
? |
mnoho
tkání |
(podle Nishizuka: FASEB J. 9, 484-496,1995)