Introduction
Biological psychiatry occupies itself by mental disorders from biological, chemical and physical point of view. The base of biological psychiatry is the assumption that human mind is connected with human body so that mental disorders (processes) are accompanied with biochemical changes, which can be measured. The second issue of biological psychiatry is causality of all natural processes, including human mind. This causality is very complexive and facts described by genetics, neuroendocrinology, immunology, biochemistry, physics etc. are included.
Biological psychiatry studies disorders in human mind from the neurochemical, neuroendocrine and genetic point of view mainly. It is postulated that changes in brain signal transmission are essential in development of mental disorders.
The purpose of this teaching material is to describe cellular and molecular properties of neurons, mechanisms of psychotropic drugs action and biological basis of mental disorders. Familiarity with biochemistry and physiology is supposed.
Apportionable parts of this teaching material are dedicated to essence of neurobiology and neurophysiology (neurons, synapses), neurochemistry (neurotransmitters, neurotrophins and receptors), psychopharmacology and biological hypothesis of schizophrenia or affective disorders.
1. NEUROBIOLOGY
The brain consists of neurons and glia. There are many different kinds of neurons and several classes of glial cells. The glia outnumber the neurons ten times or more.
1.1. Neuron
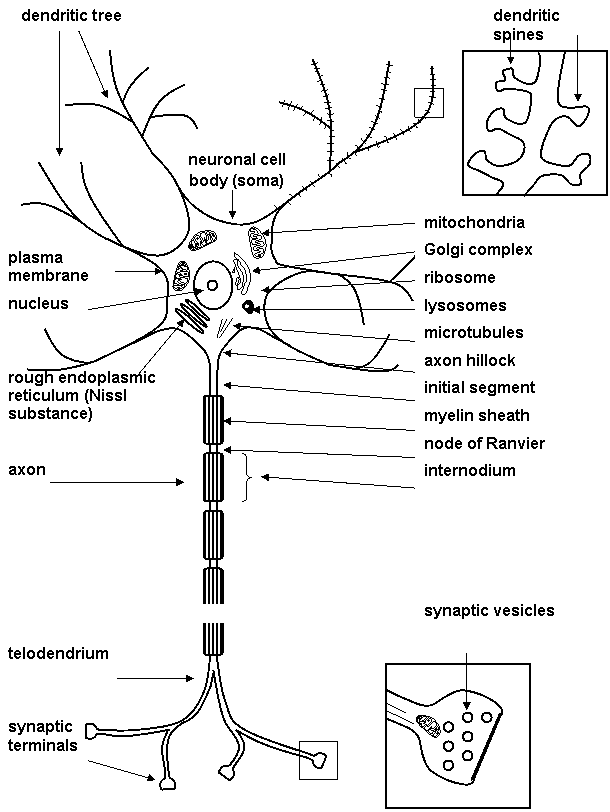
The neurons are the brain cells that are responsible for intracellular and intercellular signalling. Neurons (Fig. 1.1) share many features in common with other cells, but they are distinguished by their highly asymmetric shapes and by existence of dendrite (whose function is reception of signals from the other neurons) and axon (which is specialized for intracellular transmission of action potential from cell body to synapses). Action potential is large and rapidly reversible fluctuation in the membrane potential, that propagate along the axon. At the end of axon there are many nerve endings (or synaptic terminals, presynaptic parts, synaptic buttons, knobs). Nerve ending form an integral parts of synapse. Synapse mediates the signal transmission from one neuron to another. The dendritic tree is in continual flux and revises its synaptic connections in the course of life.
Cytoskeleton is important part of neuron. Cytoskeleton is heterogeneous network of filamentous structures: major components are microfilaments, neurofilaments and microtubules. The neuronal cytoskeleton is essential for establishing this cell shape and for axonal transport (for moving vesicles and other organelles to regions remote from the neuronal cell body). Protein called molecular motors makes use of the energy released by hydrolysis of ATP to drive axonal transport.
Figure 1.1. Neurone
1.2. Glia
Glial cells (Fig. 1.2) are:
- astrocytes
- oligodendrocytes (forming the myelin sheath)
- microglia
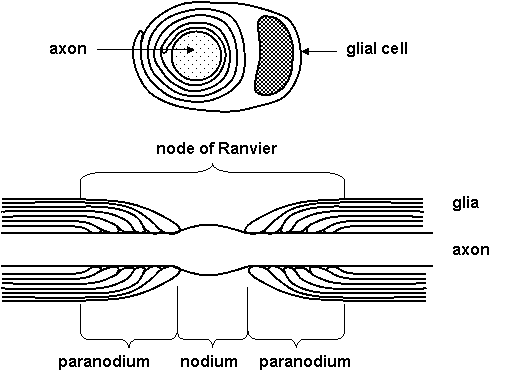
Glia are known to participate in many neuronal functions (Table 1.1). Myelination is one role of glia that is well understood. The myelin sheath surrounds many, but not all, axons in nervous system. It is formed by oligodendrocytes on central nervous system axons and by Schwann cells in the peripheral nervous system. On Figure 1.3 you can see cross section with an electron microscope - axon is surrounded by concentric circles of glial membrane. A single Schwann cell may occupy up to about 1 mm of the length of axon. There are gaps between adjacent Schwann cells of several micrometers, known as nodes of Ranvier. Region between the nodes of Ranvier is known as internode.
Table 1.1. Functions of glia
|
1. act as electrical insulator between neurons |
|
2. participate in the uptake and metabolism of the neurotransmitters |
|
3. act as a scaffolding for neuronal migration and axon outgrowth |
|
4. take up and buffer ions from the extracellular environment |
|
5. act as scavengers to remove debris produced by dying neurons |
|
6. segregate groups of neurons one from another |
|
7. provide structural support for neurons |
|
8. play a role in information handling and memory storage |
Figure 1.2. Glial cells

Figure 1.3. Myelin sheath
1.3. Plasma membrane
All cells, including neurons and glia, are enclosed by a plasma membrane.
Plasma membrane acts as:
- A barrier preventing the content of the cell from mixing with those of the extracellular space
- Effective electrical insulator
- Regulator of transport of charged and no charged molecules in and out of the cell
Plasma membranes consist of (Figure 1.4):
- Lipid bilayer (phospholipids, glycolipids, sphingomyelin, cholesterol)
- Integral and peripheral proteins (ion channels, receptors, enzymes, transporters)
Main properties of lipid bilayer are included in model of fluid mosaic:
1. Heterogeneity in the plane horizontal and vertical
2. Fluid state of most lipids at physiological conditions
3. Translation and rotation movement of membrane molecules
Dynamic properties of biological membranes enable movement and mutual interactions of membrane proteins (Figure 1.5), so membrane lipids can participate on signal transduction.
Figure 1.4. Model of plasma membrane

Figure 1.5. Lipid bilayer and function of membrane proteins
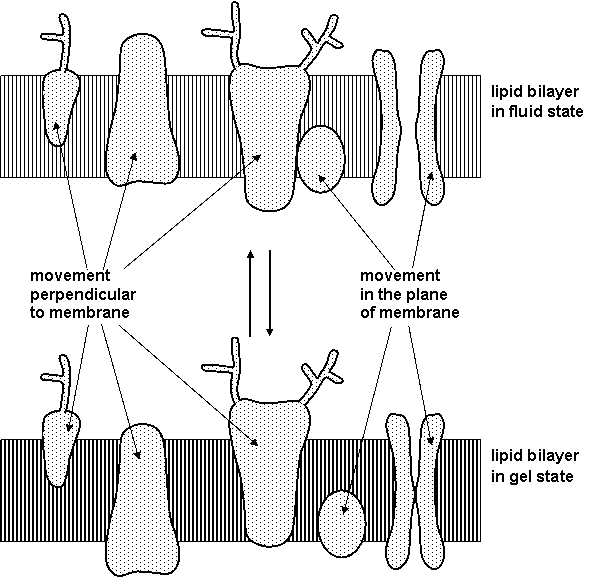
(See: Shinitzky M.: Membrane fluidity and receptor function. In: Membrane Fluidity, M. Kates, L.A. Manson, eds., Plenum Publ. Corp. 1984, pp. 585-601.)
1.4. Action potential
All living cell, including neurons, exhibit a voltage difference across plasma membrane known as membrane potential. In neurons the resting potential is usually in the range -40 to -90 mV. Information is carried from one part of the neuron to another in the form of active response of membrane termed as action potential. Action potential is a large and rapidly reversible fluctuation in the membrane potential, that propagate along the axon. Generation and propagation of action potentials are enabled by membrane ion channels.
Specific properties of axonal membranes allowing signal transmission in the form of action potentials:
- There is a threshold for generation of action potential; i.e. small random fluctuations in the membrane potential are not interpreted as useful information.
- The all-or-none law ensures full size of action potential.
- There is frequency coding of the strength and latency of the initial stimulus.
- Properties of the axonal membrane allow passive spread of the action potential depolarization along the axon, which is essential in propagation of active response. Saltatory conduction, i.e. jumping of action potentials along myelinated axons, is enabled by passive spread of depolarizing stimulus and by accumulation of ion channels in the nodes of Ranvier.
When the depolarization exceeds threshold the active response of the membrane occurs and a large change in membrane potential is observed several milliseconds in duration (Figure 1.6). The amplitude of the action potential is independent on stimulus intensity. The membrane potential depolarises very rapidly, and then there is a slightly less rapid return to the resting level. It is impossible to evoke another action potential immediately after firing of an action potential; this absolute refractory period is followed by relative refractory period, during which initial depolarization must be larger to evoke action potential.
Amplitude of action potential carries little information about stimulus that triggered them; in fact information about stimulus strength and latency can be coded in the frequency of action potential firing (Figure 1.7).
Figure 1.6. Action potential

Figure 1.7. Frequency coding
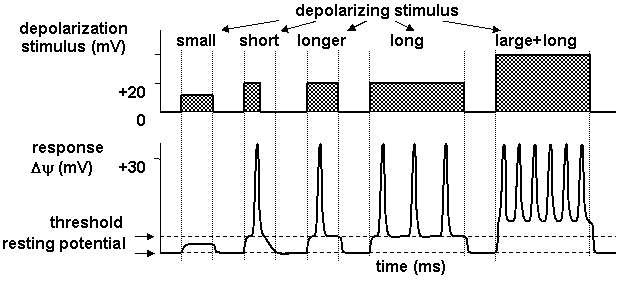
(See: Levitan I.B. a Kaczmarek L.K.: The Neuron. Oxford Univ. Press, New York, Oxford, 1997; Ganong W.F.: Přehled lékařské fyziologie. Nakl. a vyd. H&H, 1995.)
1.5. Ion channels
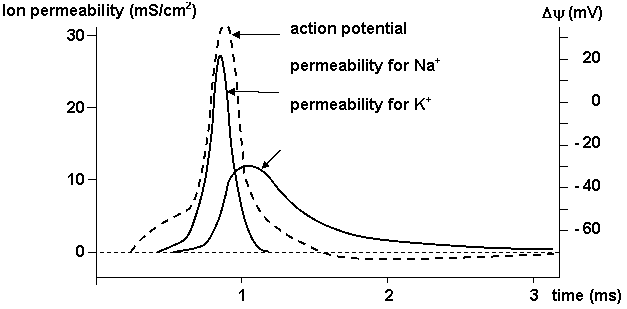
Ion channels underlie electrical signalling in neurons. It is the sum of the various currents flowing at any point that determines the neuron's membrane potential (Figure 1.8). The activities of the sodium and potassium channels responsible for axonal action potentials are dependent on voltage.
Sequence of events during the nerve impulse in a large unmyelinated axon (Figure 1.9):
- Membrane at rest
- Initial stimulus ® sodium channel opens ® Na+ enters axon ® membrane depolarisation
- At peak of impulse ® sodium channel inactivates, potassium channel opens ® K+ leaves axon ® refractory period
- After refractory period ® sodium channel inactivation removed ® both channels closed
It is now evident that there is a considerable diversity of ion currents in axons, cell bodies and dendrites. Voltage clamp and patch clamp techniques make possible revealing of heterogeneity of voltage dependent sodium, calcium, and potassium channels.
Figure 1.8. Membrane permeability for Na+ and K+ during action potential
(see Voet D., Voetová J.G.: Biochemie. Victoria Publ., Praha, 1995.)
Figure 1.9. Ion flow during action potential

(See: Atwood H.L., MacKay W.A.: Essentials of Neurochemistry. B.C. Decker Inc., Toronto, Philadelphia, 1989.)
1.6. Synapse
Neurons communicate with one another by direct electrical coupling or by the secretion of neurotransmitters (Table 1.2).
Table 1.2. Intercellular communication
|
|
|
Electrical coupling is mediated by connexins, i.e. proteins that form pores linking the cytoplasm of adjacent cells, so ions and small molecules can diffuse through pores from one cell to another.
Neurosecretion is a more complex process. Stimulation of cells is followed by an elevation of intracellular calcium, which is important intracellular messenger. Increased presynaptic Ca2+ levels allow the vesicles to fuse with the plasma membrane and to release their contents in the synaptic cleft.
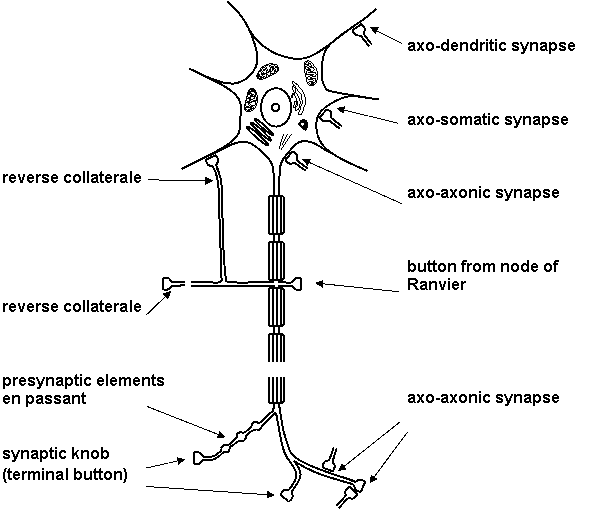
Synapses are specialized structures for signal transduction from one neuron to other. Chemical synapses are studied in the biological psychiatry. Morphology of chemical synapse is shown on Figure 1.10. Different kinds of synapses are shown on Figure 1.11.
Figure 1.10. Morphology of chemical synapse

(See: Němeček S. a kol.: Neurobiologie, Avicenum, Praha 1972)
Figure 1.11. Synapses
(See: Němeček S. a kol.: Neurobiologie, Avicenum, Praha 1972; modified.)
1.7. Chemical synapse
The main interest of biological psychiatry is knowledge of changes in signal transduction through chemical synapse (Figure 1.12) in the mental disorder and during its treatment. It is known that many psychotropic drugs act on the level of chemical synapse; it is on the level of changes of availability of neurotransmitters, regulation of receptors and adaptation of post receptor events. Therefore this teaching material is dedicated to molecular mechanisms of nerve signal transduction.
Table 1.3. Basic steps in signal transduction
|
|
|
|
Release of neurotransmitters is closely linked to the entry of calcium. With repetitive stimulation, phenomenon as facilitation, potentiation and depression of neurotransmitter release may occur.
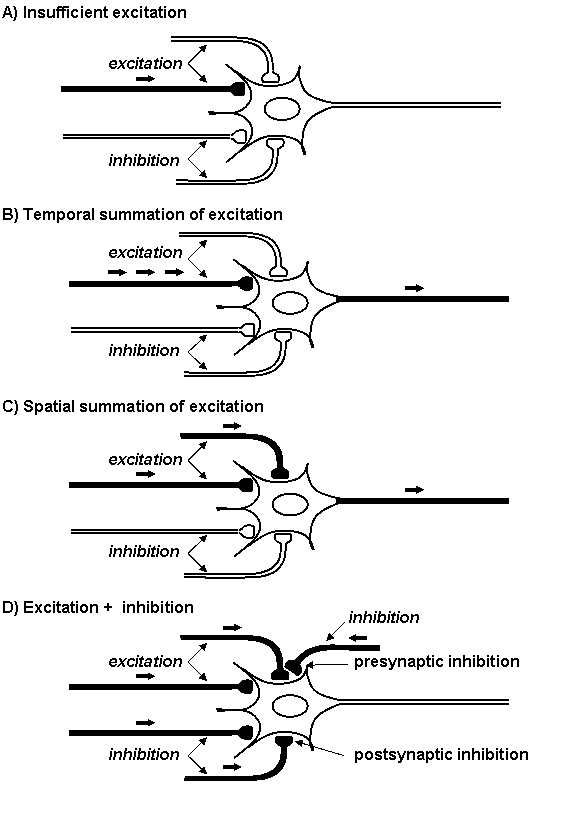
Following neurotransmitter release postsynaptic or presynaptic membrane receptors are activated. In inhibition synapse efflux of potassium ions causes hyperpolarization of postsynaptic membrane; in excitation synapse influx of sodium ions causes depolarization of postsynaptic membrane (Figure 1.13).
Temporal and/or spatial summation of synaptic potentials is necessary to the evocation of desirable physiological effect since the amount of neurotransmitters released from one vesicle is generally insufficient (Figure 1.14).
Figure 1.12. Chemical synapse - signal transduction
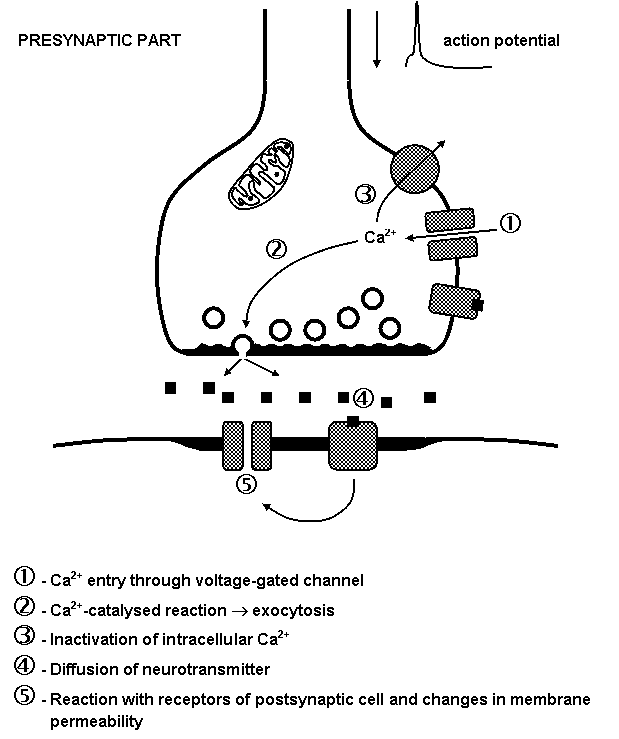
Figure 1.13. Excitation and inhibition postsynaptic potentials
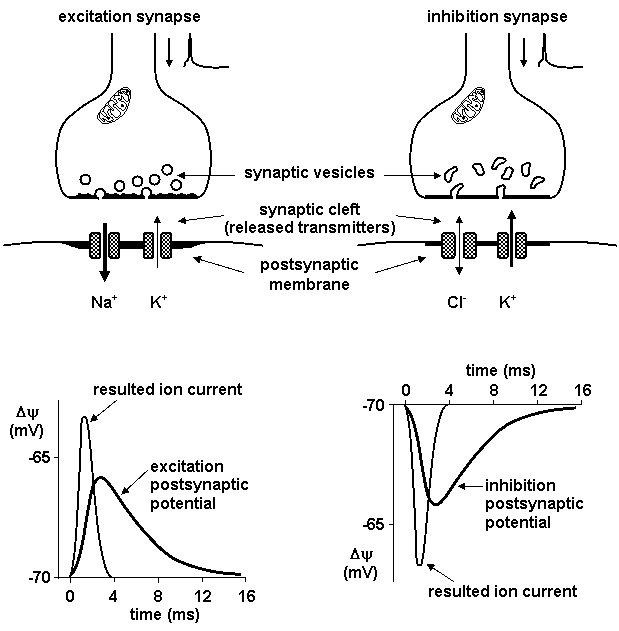
(See: Netter F.H.: Physiology and Functional Neuroanatomy. Nervous System I. CIBA-GEIGY Ltd., Basle, 1989.)
Figure 1.14. Summation of excitation and inhibition