3. Experimenty
V této kapitole jsou uvedeny protokoly vybraných experimentů využívajících fluorescenční spektroskopii. Kromě vlastního fluorescenčního měření jsou popsány i některé další metody související s popisovaným experimentem (především přípravou vzorků a zpracováním výsledků). Všechna měření lze provést na běžném spektrofluorimetru vybaveném polarizačním zařízením, ale předpokládají znalost základních biochemických a biofyzikálních postupů. Prezentované výsledky byly získány na spektrofluorimetru FluoroMax-3 (Jobin Yvon, Horiba) s programovým vybavením DATAMAX a GRAMS/32.

Nehledě na typ fluorimetru, je pro citlivost a přesnost měření důležitá čistota fluorimetrických kyvet. Pro skleněné a křemenné kyvety je doporučen tento způsob mytí:
- ponechat alespoň 24 hod namočené v kyselině chromsírové
- omýt deionizovanou vodou
- namočit na 5 hodin (ne déle!!!) do čistícího roztoku (20 tablet KOH/100 ml metanolu)
- omýt deionizovanou vodou
- namočit do koncentrované kyseliny dusičné až do použití, před nímž se kyvety vypláchnou deionizovanou vodou
3.1. Polarita rozpouštědla
V tomto demonstračním experimentu je ukázán vliv polarity rozpouštědla na fluorescenci sondy kyseliny 6-(p-toluidino)-2-naftalénsulfonové (TNS) rozpuštěné ve stejné koncentraci v roztocích obsahujících různé procento etanolu.
Vlastnosti fluorescenční
sondy kyseliny 6-(p-toluidino)-2-naftalénsulfonové (TNS):
- MW = 313,88
- rozpouštědlo pro zásobní roztok: dimetylformamid (DMF)
- rozpouštědlo pro spektroskopická měření: metanol (MeOH)
- dlouhovlnné absorpční maximum v metanolu: lexmax = 318 nm (molární extinkční koeficient: 26000 cm-1M-1)
- fluorescenční emisní maximum v metanolu: lemmax = 443 nm
- kvantový výtěžek fluorescence je závislý na okolním prostředí a je zvláště citlivý na přítomnost vody; emise je závislá na rozpouštědle
Materiál:
- kyselina 6-(p-toluidino)-2-naftalénsulfonová (TNS, Fluka)
- etanol pro UV spektroskopii (Lachema)
- destilovaná voda
- skleněné zkumavky
- mikropipety
Postup:
1. Připravte zásobní roztok TNS: odvažte cca 3 mg TNS a rozpusťte nejprve v 96% etanolu a poté upravte na koncentraci 2 mmol/l v 80% etanolu.
2. Připravte vzorky dle Tabulky 3.1, tak aby výsledná koncentrace TNS byla 5 mmol/l.
3. Změřte excitační spektra vzorků při lem = 440 nm
4. Změřte emisní spektra vzorků při lex = 320 nm
Tabulka 3.1
|
č. vzorku |
dest. voda (ml) |
etanol (80%) (ml) |
TNS (2 mmol/l) (ml) |
výsledné % etanolu |
|
|
1 |
3990 |
0 |
10 |
0,2 |
|
|
2 |
3500 |
490 |
10 |
10 |
|
|
3 |
3000 |
990 |
10 |
20 |
|
|
4 |
2000 |
1990 |
10 |
40 |
|
|
5 |
1000 |
2990 |
10 |
60 |
|
|
6 |
0 |
3990 |
10 |
80 |
|
Výsledky:
Změřená excitační a emisní spektra sondy TNS jsou na obr. 3.1 a 3.2. Měření byla provedena na spektrofluorimetru FluoroMax-3.
Obr. 3.1 Excitační spektra TNS (lem = 440 nm) v roztocích obsahujících 0,2 – 80% etanolu.
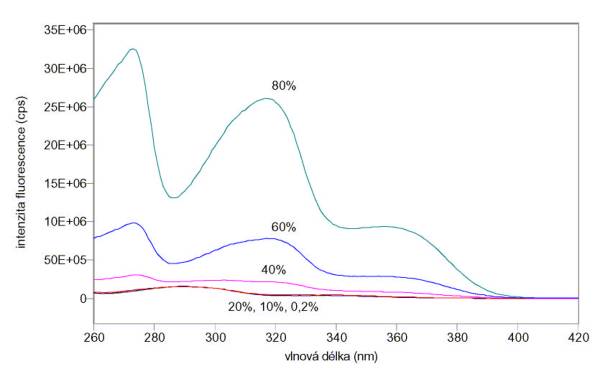
Obr. 3.2 Emisní spektra TNS (lex = 320 nm) v roztocích obsahujících 0,2 – 80% etanolu.
3.2. Vazba sond na bílkoviny
V tomto demonstračním experimentu je ukázán vliv vazby sondy ANS na albumin na její fluorescenci a zhášení fluorescence jódem.
Vlastnosti
fluorescenční sondy 1-anilinonaftalén-8-sulfonátu sodného (ANS):
- MW = 321,33
- rozpouštědlo pro zásobní roztok: dimetylformamid (DMF)
- rozpouštědlo pro spektroskopická měření: metanol (MeOH)
- dlouhovlnné absorpční maximum v metanolu: lexmax = 372 nm (molární extinkční koeficient: 7800 cm-1M-1)
- fluorescenční emisní maximum v metanolu: lemmax = 480 nm
- kvantový výtěžek fluorescence je závislý na okolním prostředí a je zvláště citlivý na přítomnost vody; emise je závislá na rozpouštědle
- podrobný popis vlastností ANS lze nalézt v práci [Slavík J.: Anilinonaphthalene sulfonate as a probe of membrane composition and function. Biochim. Biophys. Acta 694, 1-25 (1982)]
Obr. 3.3 Fluorescenční spektra sondy ANS v etanolu
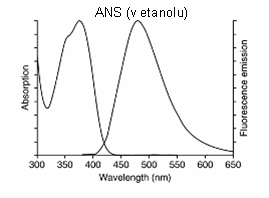
(Podle: Handbook of
Fluorescent Probes and Research Products.)
Materiál:
- 1-anilinonaftalén-8-sulfonát sodný (ANS, ICN Pharmaceuticals)
- lidský albumin (MW = 69000)
- jodid draselný (MW = 166,00)
- skleněné zkumavky
- mikropipety
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4)
Postup:
1. Připravte zásobní roztoky 2,5 mmol/l ANS, 0,25 mmol/l albuminu a 5 mol/l KJ.
2. Připravte vzorky dle Tabulky 3.2, tak aby výsledné koncentrace ANS a albuminu byly 5 mmol/l a výsledné koncentrace KJ 20 až 100 mmol/l
3. Změřte excitační a spektra vzorků při lem = 470 nm
4. Změřte emisní spektra vzorků při lex = 370 nm
Tabulka 3.2
|
č. vzorku |
pufr A (ml) |
albumin (ml) |
ANS (ml) |
KJ (ml) |
výsledná koncentrace KJ (mmol/l) |
|
0 |
4900 |
100 |
0 |
0 |
0 (vlastní fluorescence albuminu) |
|
1 |
4890 |
100 |
10 |
0 |
0 |
|
2 |
4870 |
100 |
10 |
20 |
20 |
|
3 |
4850 |
100 |
10 |
40 |
40 |
|
4 |
4830 |
100 |
10 |
60 |
60 |
|
5 |
4810 |
100 |
10 |
80 |
80 |
|
6 |
4790 |
100 |
10 |
100 |
100 |
|
7 |
4740 |
100 |
10 |
150 |
150 |
|
8 |
4690 |
100 |
10 |
200 |
200 |
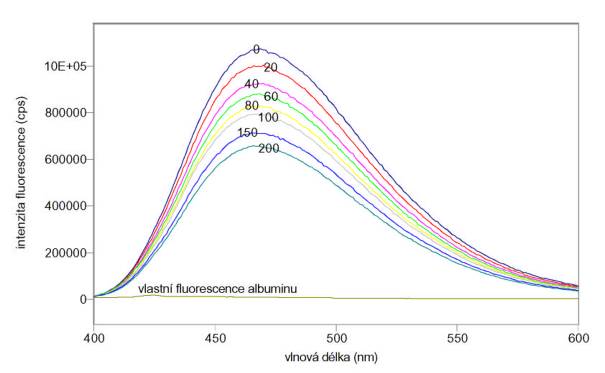
Výsledky:
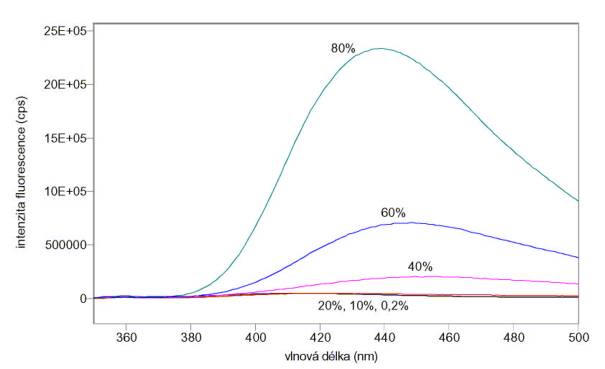
Změřená excitační a emisní spektra sondy ANS vázané k albuminu v přítomnosti různých koncentrací jodidu draselného jako zhášedla jsou na obr. 3.4 a 3.5. Měření byla provedena na spektrofluorimetru FluoroMax-3.
Obr. 3.4 Excitační spektra ANS (lem = 470 nm) vázaného na albumin v roztocích obsahujících 0 – 200 mmol/l jodidu draselného.
Obr. 3.5 Emisní spektra ANS (lex = 370 nm) vázaného na albumin v roztocích obsahujících 0 – 200 mmol/l jodidu draselného.
3.3. Dynamický stav lipidových membrán
V tomto experimentu jsou ukázány možnosti měření polarizované fluorescence v ustáleném stavu (tj. při kontinuálním buzení). Jsou testovány možnosti fluorescenční membránové sondy 1,6-difenyl-1,3,5-hexatrienu (DPH) pro stanovení změn dynamického stavu lipidových dvojvrstev. Měření je provedeno jednak v modelových membránách, liposomech, jednak v erytrocytárních membránách. Je proto uveden také postup přípravy velkých jednovrstevných liposomů a erytrocytárních ghostů.
Vlastnosti
fluorescenční sondy 1,6-difenyl-1,3,5-hexatrienu (DPH):
- MW = 232,32
- rozpouštědlo pro zásobní roztok: dimetylformamid (DMF), acetonitril (MeCN)
- rozpouštědlo pro spektroskopická měření: metanol (MeOH), aceton
- zásobní roztoky DPH se často připravují v tetrahydrofuranu (THF); dlouhodobé skladování v THF se však nedoporučuje kvůli možné tvorbě peroxidů v tomto rozpouštědle
- dlouhovlnné absorpční maximum v metanolu: lexmax = 350 nm (molární extinkční koeficient: 88000 cm-1M-1)
- fluorescenční emisní maximum v metanolu: lemmax = 452 nm
- DPH a jeho deriváty prakticky nefluoreskují ve vodě. Absorpční a emisní spektra mají více píků; vlnová délka, rozlišení a relativní intenzita těchto píků závisí na prostředí. Uvedené hodnoty lexmax a lemmax jsou pro nejintenzivnější pík v metanolu.
Obr. 3.6 Lokalizace fluorescenčních sond v lipidové dvojvrstvě
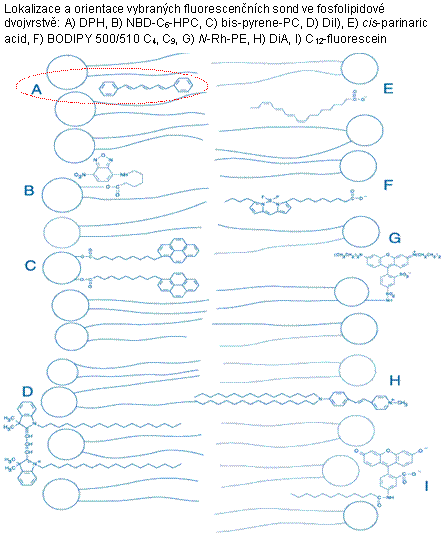
(Podle: Handbook of
Fluorescent Probes and Research Products.)
Materiál:
- 1,6-difenyl-1,3,5-hexatrien (DPH, Sigma)
- fosfatidylcholin izolovaný z hovězího mozku
- ACD roztok (0,8% kyselina citrónová, 2,2% citrát sodný (Na3C6H5O7), 2,4% glukóza, pH 4,3)
- plná krev
- centrifuga
- zařízení LiposoFast-Basic
- skleněné zkumavky
- mikropipety
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4)
- hypotonický pufr pro hemolýzu (izotonický pufr A se 20x naředí redestilovanou vodou a pH se upraví na 7.4 pomocí NaOH a HCl)
Postup:
Příprava velkých
jednovrstevných liposomů (LUV)
Liposomy jsou váčky (vezikuly) uzavírající vodný roztok membránou tvořenou především fosfolipidy. Tvoří se spontánně, když jsou fosfolipidy dispergovány ve vodném prostředí. V závislosti na způsobu přípravy mohou vzniknout liposomy 1. mnohovrstevné (multilamellar vesicles, MLV), 2. malé jednovrstevné (small unilamellar vesicles, SUV) nebo 3. velké jednovrstevné (large unilamellar vesicles, LUV). V našich experimentech jsme použili LUV o průměru cca 100 nm.
Obr. 3.7 Tvorba lipidových dvojvrstev a liposomů

Liposomy byly připravovány z fosfatidylcholinu (PC) izolovaného z bílé hmoty hovězího mozku. PC byl separován z hrubého extraktu lipidů sloupcovou chromatografií. Výsledná čistota fosfolipidů byla lepší než 95% (určeno dvourozměrnou tenkovrstevnou chromatografií). Fosfolipidy byly uchovávány v dusíkové atmosféře ve směsi chloroform/metanol (2 : 1, v/v) při -30 °C. Koncentrace fosfolipidů byla určena z měření koncentrace fosforu (viz níže). Nejprve byly připraveny velké mnohovrstevné liposomy (modifikovanou Banghamovou metodou) a z nich byly protlačováním přes polykarbonátové filtry pomocí zařízení Liposofast-Basic (Avestin, Inc.) připraveny LUV:
1.
přenést 1 až 20 mg fosfolipidu ve směsi
chloroform/metanol (2:1) do skleněné nádobky a rozpouštědlo odpařit proudem
dusíku při teplotě kolem
2. nádobku umístit po několik hodin ve vakuu, aby byly odstraněny zbytky rozpouštědla
3. přidat 2 ml pufru A a vzorek inkubovat v 50 °C po dobu 5 minut
4. intenzivně třepat a nakonec krátce sonikovat (cca 5 s) v sonikátoru XL 2020 (Heat Systems - Ultrasonics) na stupni 4
5. inkubovat vzorek v 50 °C po dobu 30 minut a poté jej krátce protřepat a naředit pufrem A na požadovanou koncentraci fosfolipidu (např. 1 mmol PC/ml); tím jsou připraveny PC-MLV
6. zařízení Liposofast-Basic vyčistit etanolem
7. první polovinu držáku filtrů zasunout do krytu a vložit polykarbonátový filtr s póry 100 nm na „O“ ring
8. přidat druhou část držáku filtrů do krytu tak, aby „O“ ringy byly vidět pozorovacím otvorem
9. utáhnout ručně pevně tak abychom se vyhnuli zkroucení držáku a aby mrtvý objem byl co nejmenší (filtry jsou těsně stlačeny mezi „O“ ringy)
10. zatlačit a současně jemně zašroubovat speciální injekční stříkačky do držáku filtrů (jedna obsahuje MLV)
11. protlačit liposomy přes filtr alespoň 11 krát (lichý počet!!!)
12. odebrat LUV a naředit
Příprava
erytrocytárních ghostů
Membrány z erytrocytů byly připraveny modifikovanou metodou dle Dodge et al. [Dodge J.T., Mitchell C., Hanahan D.J.: The preparation and chemical characteristics of hemoglobin-free ghosts of human erythrocytes. Arch. Biochem. Biophys. 100, 119‑130 (1963)]. Všechny operace se provádějí při 4°C:
1. krev (cca 4 ml) odebrat do ACD roztoku (0,5 ml) a centrifugovat v odběrové zkumavce při 200g (1000 až 1200 RPM na JK23, výkyvný rotor), 20oC, 15 min.
2. odebrat opatrně plazmu obohacenou trombocyty (PRP) i s horní vrstvičkou sedimentu (erymasy) a zbytek erymasy 3 x promýt pufrem A (4oC, 10 min, 1000 g = 3000 RPM na K23)
3.
sediment z původních 4 ml krve se rozpipetuje do
dvou 50 ml zkumavek, přidá se cca 30 ml hypotonického pufru, pečlivě se
promíchá a potom centrifuguje (20000 g,
4. ghosty se ještě alespoň 2x promyjí hypotonickým pufrem (poměr buňky:promývací roztok je asi 1:30); pečlivé odstraňování supernatantu je velmi důležité
5. po posledním točení se membrány roztřepou v cca 2 ml pufru A a převedou se do ependorfek v objemu odpovídajícím původnímu objemu krve
6. odebere se vzorek (20 ml) pro stanovení proteinů a fosforu a membrány se dají zamrazit.
Stanovení fosforu v
membránách
Fosfát je určován metodou dle [Bartlett, G.R.: Phosphorus assay in column chromatography. J. Biol. Chem. 234 (1959) 466-468; Wagner, H., Lissau, A., Holzi, J., Horammer, L.: The incorporation of 32P into inositolphosphatides of the rat brain. J. Lipid Res. 3 (1962) 177-180].
Roztoky:
- 2,5% molybdenan amonný (25 g/l)
- 0,02 M KH2PO4 (272,2 mg/100 ml)
- 70% HClO4
- 10% kyselina askorbová (vždy čerstvá)
- standard: 100x zředěný roztok 0,02 mol/l KH2PO4 (1ml+99 ml vody)
Postup:
1. vzorky odpařit pod dusíkem v kalibrovaných zkumavkách
2. vzorek, standard (S1 = 0,25 ml 0,2 mmol/l KH2PO4; S2 = 0,5 ml 0,2 mmol/l KH2PO4) nebo slepý vzorek + 0,30 ml HClO4 (70%)
3.
spalovat v pískové lázni 60 min při
4. po vychladnutí doplnit do 2,0 ml redestilovanou vodou, třepat
5. přidat 0,25 ml molybdenanu amonného, třepat
6. přidat 0,25 ml kyseliny askorbové, třepat (výsledný objem je 2,5 ml)
7. 5 min vařit ve vodní lázni
8. 5 min chladit v ledové vodě
9. změřit absorbanci Avzorek při 820 nm proti slepému vzorku
10. provést výpočet na základě skutečnosti, že ve standardu S1 je 0,05 mmol fosfátu (Pi), ve standardu S2 je 0,10 mmol Pi:
· kalibrační faktor f pro 0,05 mmol Pi je:
(průměr z S1 + (průměr z S2/2))/2 = f
· koncentrace Pi ve vzorku je:
c(mmol Pi/ml) = c(mmol/l) = (0,05 . Avzorek)/(f . V)
kde je V – objem vzorku odebraného pro měření Pi v mililitrech
· přibližnou koncentraci fosfolipidů (MW @ 752) v mg/ml dostaneme vydělením této hodnoty c(mmol/l) koeficientem 1,33
Metoda
fluorescenčních sond
Pro měření relativních změn fluidity plazmatických membrán byla použita metoda fluorescenčních sond s hydrofobní membránovou sondou 1,6-difenyl-1,3,5-hexatrien (DPH, Sigma). DPH byl rozpuštěn v acetonu v koncentraci 6 mmol/l a tento zásobní roztok byl uložen v mrazničce. Pro měření byl připraven vždy čerstvý roztok v pufru A v koncentraci 6 µM DPH (injektováním pomocí Hamiltonovy stříkačky 10 ml za intenzivního míchání). Tento roztok byl smíchány s pufrem A a suspenzí liposomů nebo membrán tak, aby výsledná koncentrace DPH byla 2 µM a koncentrace fosfolipidů cca 100 µmol/l v případě buněčných membrán a 50 mmol/l v případě PC-LUV. Po 60 minutách inkubace v 37°C byla měřena polarizovaná fluorescence (anizotropie fluorescence) při excitační vlnové délce 350 nm a emisní vlnové délce 450 nm na spektrofluorimetru FluoroMax-3 (Jobin Yvon, Horiba) s polarizačním příslušenstvím. Měření daného vzorku bylo opakováno alespoň 10x a hodnoty byly zprůměrňovány.
Polarizovaná fluorescence roztoků se měří ve směru kolmém ke směru
budícího paprsku, který je polarizován ve svislém směru. Je přitom nutno
provádět korekci na vliv emisního monochromátoru spektrofluorimetru na
polarizaci jím procházejícího záření. Anizotropie
fluorescence se určuje ze vztahu
(1.21) r
= (IIIV - G I^V)/((IIIV + 2 G I^V)
kde IIIV a I^V jsou složky světelné intenzity rovnoběžné nebo kolmé k
směru (vertikálnímu) polarizace budícího záření a G je korekční faktor, který
lze změřit při excitačním záření polarizovaném vodorovně jako poměr I^H/IIIH.
Výsledky:
Pro ilustraci jsou v tabulce uvedeny výsledky měření
anizotropie fluorescence DPH (rDPH) v ghostech (při
Tabulka 3.3 Anizotropie fluorescence sondy DPH v ghostech
|
SIMVASTATIN |
GHOSTY |
SÉRUM |
|||||||
|
doba podávání léku snižujícího sérový cholesterol |
rDPH |
poměr
cholesterol/fosfolipidy v ghostech |
celkový sérový cholesterol |
||||||
|
0 |
0,2382 |
± |
0,0065 |
0,756 |
± |
0,137 |
6,57 |
± |
1,26 |
|
1 měsíc |
0,2338 |
± |
0,0062 |
0,804 |
± |
0,111 |
6,20 |
± |
0,89 |
|
2 měsíce |
0,2320 |
± |
0,0115 |
0,711 |
± |
0,085 |
5,10 |
± |
0,42 |
|
13-16 měsíců |
0,2316 |
± |
0,0062 |
0,788 |
± |
0,209 |
5,02 |
± |
0,39 |
|
zdravé kontroly |
0,2280 |
± |
0,0067 |
0,683 |
± |
0,127 |
3,83 až 5,8 |
||
Anizotropie fluorescence rDPH v liposomech
připravených z fosfatidylcholinu je mnohem menší, než rDPH
v ghostech. Pro ilustraci jsou v následující tabulce 3.4 uvedeny
hodnoty rDPH v PC-LUV (při
Tabulka 3.4 Anizotropie fluorescence sondy DPH ve velkých jednovrstevných liposomech
|
PC-LUV |
|
|
koncentrace imipraminu (mmol/l) |
rDPH |
|
0 |
0,08624 |
|
0,01 |
0,08513 |
|
0,1 |
0,08484 |
|
1 |
0,08484 |
|
10 |
0,08550 |
|
100 |
0,09156 |
3.4. Vlastní fluorescence bílkovin
Vlastní fluorescenci proteinů je způsobena aromatickými aminokyselinami v nich obsaženými: tryptofanem (Trp), tyrozinem (Tyr) a fenylalaninem (Phe). Dominující je fluorescence z tryptofanu, naopak prakticky vůbec se neuplatňuje fenylalanin (viz kap. 2.1). V tomto experimentu jsou změřena fluorescenční excitační a emisní spektra albuminu a čistého tryptofanu, tyrozinu a fenylalaninu.
Vlastnosti L-tryptofanu:
- MW = 204,23
- absorpční maximum: lexmax = 295 nm
- fluorescenční emisní maximum: lemmax = 353 nm
- emise Trp je vysoce závislá na polaritě a okolním prostředí
Vlastnosti
L-tyrozinu:
- MW = 181,19
- absorpční maximum: lexmax = 275 nm
- fluorescenční emisní maximum: lemmax = 304 nm
- emise Tyr je relativně málo citlivá na polaritu rozpouštědla
Vlastnosti
L-fenylalaninu:
- MW = 165,19
- absorpční maximum: lexmax = 260 nm
- fluorescenční emisní maximum: lemmax = 282 nm
- emise Phe je strukturovaná
Fluorescence proteinů je obvykle excitována při 280 nm nebo při delších vlnových délkách, takže fenylalanin není ve většině experimentů excitován. Navíc je kvantový výtěžek fluorescence Phe velmi malý (kolem 0,02). Tryptofanovou fluorescenci v proteinech lze selektivně excitovat při 295-305 nm.
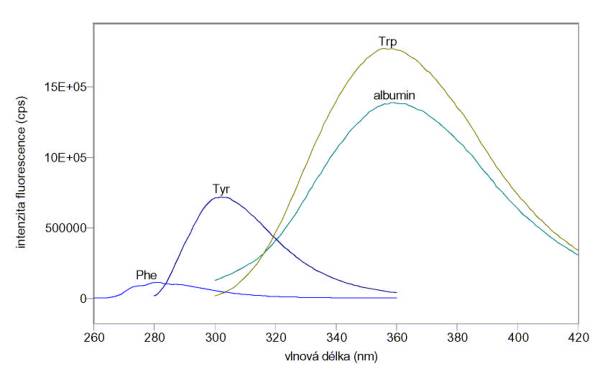
Obr. 3.8 Fluorescenční spektra tryptofanu (Trp), tyrozinu (Tyr) a fenylalaninu (Phe)
![]()
![]()
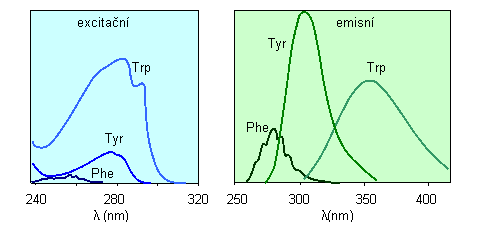
(Podle: Lakowicz
1999.)
Materiál:
- L-tryptofan (Aldrich), L-tyrozin (Fluka), L-fenylalanin (Fluka)
- lidský albumin (MW = 69000)
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4)
- skleněné zkumavky, mikropipety, …
Postup:
- Připravte zásobní roztoky 10 mmol/l Trp, 10 mmol/l Tyr, 10 mmol/l Phe a 0,25 mmol/l albuminu v pufru A.
- Pro vlastní měření je nařeďte pufrem A na koncentrace 20 mmol/l Trp, 100 mmol/l Tyr, 10 mmol/l Phe a 5 mmol/l albuminu.
- Změřte excitační a emisní spektra vzorků za podmínek uvedených v tabulce3.5
Tabulka 3.5
|
|
excitační spektrum |
emisní spektrum |
||
|
lem (nm) |
lex (nm) |
lex (nm) |
lem (nm) |
|
|
L-tryptofan |
350 |
240-320 |
280 |
300-420 |
|
L-tyrozin |
300 |
240-290 |
270 |
280-360 |
|
L-fenylalanin |
280 |
240-270 |
250 |
260-360 |
|
albumin |
350 |
240-320 |
280 |
300-420 |
Výsledky:
![]() Změřená excitační a emisní spektra
vlastní fluorescence tryptofanu (Trp), tyrozinu (Tyr), fenylalaninu (Phe) a
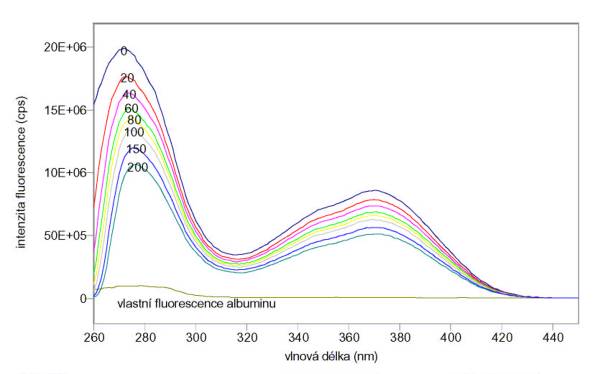
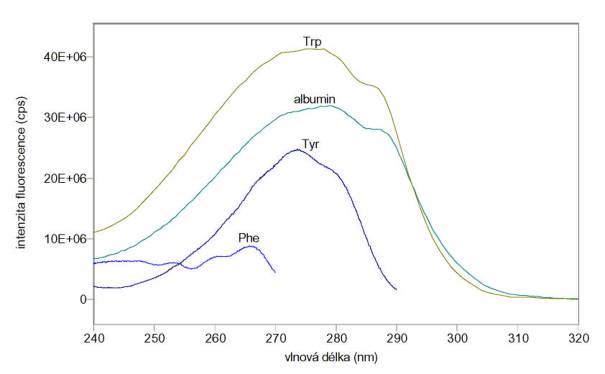
albuminu v pufru A jsou na obr. 3.9 a 3.10. Je vidět, že prakticky veškerá
vlastní fluorescence albuminu pochází od tryptofanu. Měření byla provedena na
spektrofluorimetru FluoroMax-3.
Změřená excitační a emisní spektra
vlastní fluorescence tryptofanu (Trp), tyrozinu (Tyr), fenylalaninu (Phe) a
albuminu v pufru A jsou na obr. 3.9 a 3.10. Je vidět, že prakticky veškerá
vlastní fluorescence albuminu pochází od tryptofanu. Měření byla provedena na
spektrofluorimetru FluoroMax-3.
Obr. 3.9 Excitační spektra 20 mmol/l Trp (lem = 350 nm), 100 mmol/l Tyr (lem = 300 nm), 10 mmol/l Phe (lem = 280 nm) a 5 mmol/l albuminu (lem = 350 nm).
Obr. 3.10 Emisní spektra 20 mmol/l Trp (lex = 280 nm), 100 mmol/l Tyr (lex = 270 nm), 10 mmol/l Phe (lex = 250 nm) a 5 mmol/l albuminu (lex = 280 nm).
3.5. Fluorescenční značení DNA
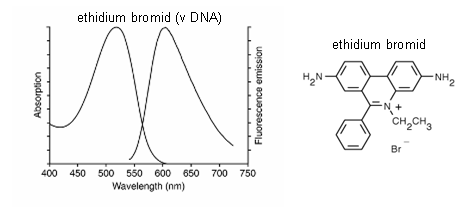
Nukleotidy a nukleové kyseliny obecně nefluoreskují. Pro jejich vizualizaci a identifikaci se proto používá řada fluorescenčních sond (viz kap. 2.2.2 a Tabulka 2.6). V tomto experimentu je ukázána možnost použití sondy ethidium bromidu pro barvení jader neživých buněk granulocytů nebo lymfocytů.
Vlastnosti sondy
ethidium bromidu:
- MW = 394,31
- dobře rozpustný ve vodě
- ve vodě fluoreskuje málo, po navázání k DNA se fluorescence zvyšuje asi 30 krát
- doba dohasínání fluorescence ve vodě je asi 1,7 ns, po vazbě k dvouřetězcové DNA se zvyšuje na 20 ns
- vazba k DNA se uskutečňuje vmezeřováním rovinného aromatického kruhu mezi páry bazí dvouřetězcové DNA
- absorpční maximum v DNA: lexmax = 523 nm
- fluorescenční emisní maximum v DNA: lemmax = 604 nm
Obr. 3.11 Fluorescenční spektra ethidium bromidu
(Podle: Handbook of
Fluorescent Probes and Research Products.)
Materiál:
- ethidium bromid (EB, Merck)
- plná krev
- dextran
- Ficoll-Paque Plus (Amersham Biosciences)
- centrifuga
- termostat
- skleněné zkumavky
- mikropipety
- fosfátový pufr (PBS, pH 7,4)
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4)
- ACD roztok (0,8% kyselina citrónová, 2,2% citrát sodný (Na3C6H5O7), 2,4% glukóza, pH 4,3)
Postup:
Izolace granulocytů z
krve
1. krev odebrat do ACD roztoku (v objemovém poměru 1:9)
2. v umělohmotné injekční stříkačce promíchat 4 díly krve a 1 díl 6% dextranu v PBS
3. nechat (stříkačku špičkou vzhůru) 30 minut v 37°C, potom vytlačit plazmu
4. promýt v PBS (centrifugace 20 min při 1500 RPM na JK23)
5. je-li
příměs erytrocytů provést hypotonní lýzu (0,25 ml vzorku + 0,75 ml vody, při
Izolace lymfocytů
z plné krve
1. krev odebrat do citrátového roztoku (nesrážlivá krev)
2. promíchat 2 ml krve a 2 ml pufru A
3. přidat 3 ml Ficoll-Paque Plus do zkumavky a opatrně navrstvit 4 ml zředěné krve
4.
centrifugovat při 400g po dobu 30 min při
5. odsát horní vrstvu (může být uchována pro pozdější použití)
6. přenést vrstvu s lymfocyty do centrifugační kyvety (cca 2 ml)
7. přidat alespoň 3 objemy pufru A, promíchat
8.
centrifugovat při 60-100g po 10 min při
9. odstranit supernatant a opakovat promytí
10. suspendovat lymfocyty v pufru A v koncentraci 5.107/ml
Barvení DNA ethidium
bromidem:
1. připravit zásobní roztok ethidium bromidu ve vodě (2,5 mmol/l)
2. smíchat pufr A, granulocyty nebo lymfocyty a ethidium bromid, tak aby výsledné koncentrace byly cca 106 buněk/ml a 10 mmol/l ethidium bromidu
3. změřit excitační fluorescenční spektrum při lem = 610 nm a emisní spektrum při lex = 520 nm
4. vzorek několikrát rychle zmrazit a rozmrazit, krátce turaxovat a znovu měřit excitační a emisní spektra
Výsledky:
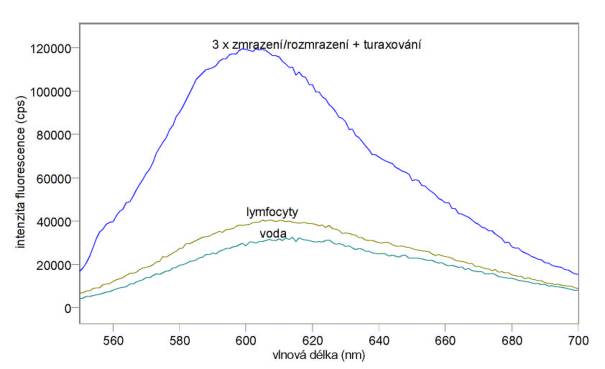
Změřená excitační a emisní spektra ethidium bromidu ve pufru A, po přidání lymfocytů a po vazbě k jejich DNA jsou uvedena na obr. 3.12 a 3.13. Měření byla provedena na spektrofluorimetru FluoroMax-3.
Obr. 3.12 Excitační fluorescenční spektra 10 mmol/l ethidium bromidu ve vodě a v DNA lymfocytů (lem = 610 nm).
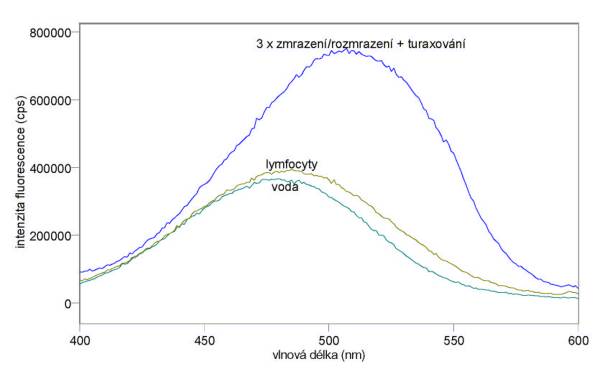
Obr. 3.13 Emisní fluorescenční spektra 10 mmol/l ethidium bromidu ve vodě a v DNA lymfocytů (lex = 520 nm).
3.6. Kalcium
Kalciové sondy jsou nejčastěji používané nitrobuněčné indikátory (viz kap. 2.3.5). Protože neprocházejí buněčnou membránou, dostávají se do buněk mikroinjekcí nebo elektroporací, nebo jako acetoxylmetylestery (AM estery), které jsou uvnitř buňky štěpeny nitrobuněčnými esterázami za vzniku negativně nabitých sond.
V tomto experimentu jsou vyzkoušeny vlastnosti dvou sond pro měření Ca2+: Fluo-3 a Quin-2. Po vazbě kalcia vykazuje Quin-2 jak změnu intenzity fluorescence, tak posuv absorpčního maxima, Fluo-3 téměř nefluoreskuje za nepřítomnosti Ca2+. Jedná se tedy o sondy, pro něž je koncentrace Ca2+ dána vztahem (viz kap. 2.3.5):
(2.1) ca = Kd (I-Imin)/(Imax-I)
kde je Kd – disociační konstanta vazebného místa pro kalcium na indikátoru, Imin – intenzita fluorescence indikátoru, když není navázán žádný vápník, Imax – intenzita fluorescence plně obsazeného indikátoru, I – intenzita fluorescence ve vzorku, kde je jen část vazebných míst indikátoru obsazena Ca2+. Měření Imax vyžaduje lýzu buněk a titraci uvolněného indikátoru nebo použití ionoforů, aby došlo k saturaci nitrobuněčného indikátoru kalciem.
Vlastnosti sondy
Quin-2:
· MW = 541,51 (pro bezvodou látku)
- mírná rozpustnost ve vodě (pH > 6)
- neprostupuje membránou, ale prodává se i AM ester (Quin-2, AM), který membránou prostupuje
- kvantový výtěžek fluorescence volné formy je 0,03; po navázání kalcia vzrůstá na 0,14
- doba dohasínání fluorescence volné formy je 1,35 ns, po vazbě Ca2+ se zvyšuje na 11,6 ns
- absorpční maximum volné formy 356 nm, vázané formy 336 nm
- fluorescenční emisní maximum volné formy 500 nm, vázané formy 503 nm
- disociační konstanta Kd = 60,0 nmol/l
Vlastnosti sondy
Fluo-3:
- MW = 769,5 (Fluo-3 tetraoctová kyselina)
- rozpustná ve vodě (pH > 6)
- neprostupuje membránou, ale prodává se i AM ester (Fluo-3, AM), který membránou prostupuje
- kvantový výtěžek fluorescence se po navázání kalcia zvýší asi 40 krát
- doba dohasínání fluorescence volné formy je 0,04 ns, po vazbě Ca2+ se zvyšuje na 1,28 ns
- absorpční maximum 504 nm
- fluorescenční emisní maximum 526 nm
- disociační konstanta Kd = 390 nmol/l
Obr. 3.14 Absorpční a fluorescenční spektrum sondy Fluo-3 saturované kalciem
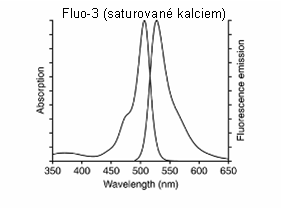
(Podle: Handbook of
Fluorescent Probes and Research Products.)
Materiál:
- Quin-2 (Sigma), Fluo-3 (Fluka)
- CaCl2 (MW = 110,99)
- EGTA
- skleněné zkumavky
- mikropipety
Postup:
1.
Quin-
2. CaCl2 rozpusťte ve vodě v koncentracích 10 mmol/l a 10 mmol/l
3. připravte 1 mmol/l EGTA ve vodě
4. smíchejte vodu, sondu a CaCl2 dle Tabulky 3.6 a změřte
§ excitační spektra Fluo-3 při lem = 540 nm; emisní spektra Fluo-3 při lex = 480 nm
§ excitační spektra Quin-2 při lem = 500 nm; emisní spektra Quin-2 při lex = 336 nm
místo měření spekter můžete měřit jen intenzitu fluorescence:
§ Fluo-3 při lex = 504 nm a lem = 526 nm
§ Quin-2 při lex = 356 nm a lem = 500 nm
Tabulka 3.6
|
voda (ml) |
Fluo-3 (ml) |
Quin-2 (ml) |
CaCl2 10 mmol/l (ml) |
CaCl2 10 mmol/l (ml) |
EGTA 1 mmol/l (ml) |
výsledná konc. Ca2+ (mmol/l) |
|
3975 |
25 |
0 |
0 |
0 |
1000 |
0 |
|
4975 |
25 |
0 |
0 |
0 |
0 |
0 |
|
4945 |
25 |
0 |
30 |
0 |
0 |
0,06 |
|
4775 |
25 |
0 |
200 |
0 |
0 |
0,4 |
|
4974 |
25 |
0 |
0 |
1 |
0 |
2 |
|
4965 |
25 |
0 |
0 |
10 |
0 |
20 |
|
4955 |
25 |
0 |
0 |
20 |
0 |
40 |
|
4925 |
25 |
0 |
0 |
50 |
0 |
100 |
|
3975 |
0 |
25 |
0 |
0 |
1000 |
0 |
|
4975 |
0 |
25 |
0 |
0 |
0 |
0 |
|
4945 |
0 |
25 |
30 |
0 |
0 |
0,06 |
|
4965 |
0 |
25 |
0 |
10 |
0 |
20 |
|
4955 |
0 |
25 |
0 |
20 |
0 |
40 |
Poznámka: Do vzorku s nulovou koncentrací přidaného CaCl2 byl přidán 0,2 mmol/l EGTA pro vychytání iontů Ca2+.
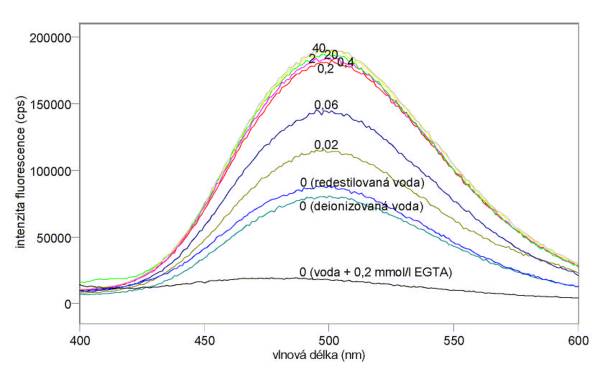
Výsledky:
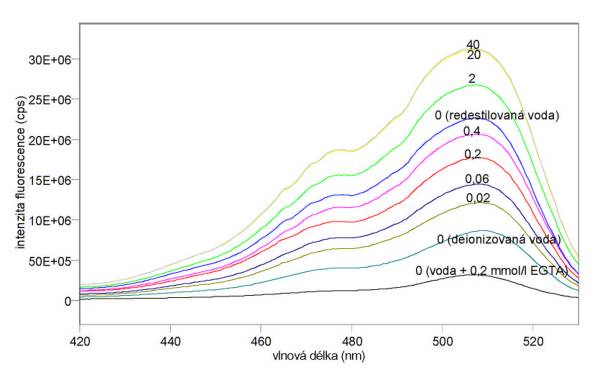
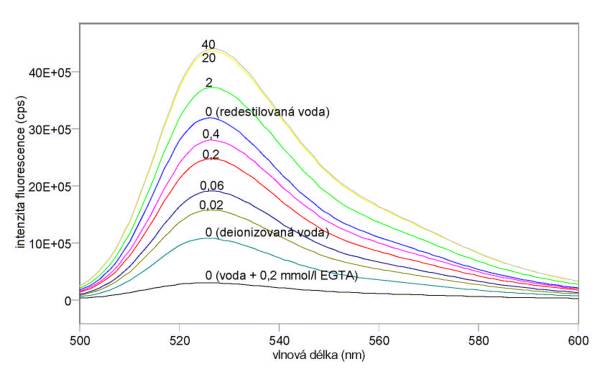
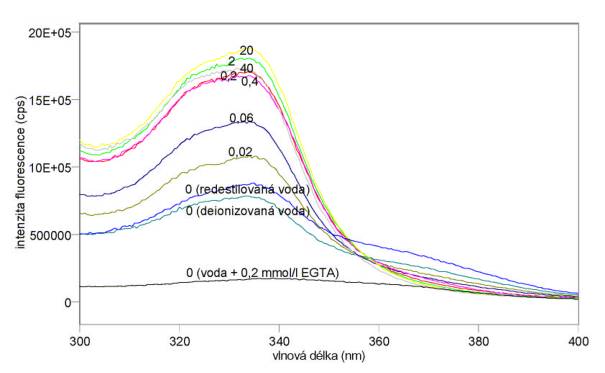
Změřená excitační a emisní spektra sondy Fluo-3 při různých koncentracích kalcia jsou uvedena na obr. 3.15 a 3.16. Excitační a emisní spektra sondy Quin-2 při různých koncentracích kalcia jsou uvedena na obr. 3.17 a 3.18. Měření byla provedena na spektrofluorimetru FluoroMax-3. Výsledky mimo jiné ukazují, že v použité redestilované vodě byly přítomny ionty Ca2+, které měření při nízkých koncentracích CaCl2 zkreslily. Je vhodnější použít deionizovanou vodu. Jak se dalo předpokládat z hodnot disociačních konstant, je sonda Quin-2 citlivější na změny koncentrací kalcia při jeho nižších koncentracích (kolem 0,06 mmol/l), zatímco sonda Fluo-3 je nejcitlivější při vyšších koncentracích Ca2+ (kolem 0,4 mmol/l).
Obr. 3.15 Excitační fluorescenční spektra 10 mmol/l Fluo-3 (lem = 540 nm) při různých koncentracích CaCl2 (mmol/l).
Obr. 3.16 Emisní fluorescenční spektra 10 mmol/l Fluo-3 (lex = 480 nm) při různých koncentracích CaCl2 (mmol/l).
Obr. 3.17 Excitační fluorescenční spektra 10 mmol/l Quin-2 (lem = 500 nm) při různých koncentracích CaCl2 (mmol/l).
Obr. 3.18 Emisní fluorescenční spektra 10 mmol/l Quin-2 (lex = 336 nm) při různých koncentracích CaCl2 (mmol/l).
3.7. Membránový potenciál
V tomto experimentu je popsán jednoduchý způsob detekce změn membránového potenciálu živých buněk pomocí fluorescenčních sond citlivých na elektrický potenciál přes membránu (viz. kap. 2.3.7). Experiment je proveden s buňkami lymfocytů izolovaných z periferní krve a potenciálovou fluorescenční sondou 3,3´-dihexyloxakarbocyanin jodidem (DiOC6(3)). Změna membránového potenciálu je indukována valinomycinem, vysokými koncentracemi mimobuněčného KCl, ouabainem a Tritonem X 100.
Triton X 100 je neiontový detergent, který rozpouští membrány a tvoří micely s membránovými lipidy a proteiny. Ouabain je inhibitor Na+K+-ATPázy. Valinomycin je ionofor přenašečového typu pro K+, tzn. zvyšuje permeabilitu membrán tím, že selektivně váže ion K+, difunduje membránou a uvolňuje K+ na druhé straně membrány (tento efekt se může uplatňovat jen do vyrovnání koncentrací iontů K+ na obou stranách membrány).
Sonda DiOC6(3) je citlivá na membránový potenciál jak plazmatické membrány, tak i mitochondriální membrány [Salvioli et al.: FEBS Letters 411, 77-82, 1997]. Pro měření změn mitochondriálního membránového potenciálu se proto musí použít specifické mitochondriální inhibitory [Kalbacová et al.: Cytometry 52A, 110-116, 2003].
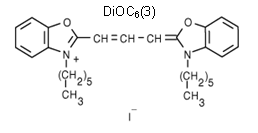
Vlastnosti sondy DiOC6(3):
- MW = 572,53
- nízká rozpustnost ve vodě
- patří do skupiny karbocyninových potenciometrických fluorescenčních sond s pomalou odezvou
- rozpouštědlo pro zásobní roztok: dimetylsulfoxid (DMSO) nebo etanol (EtOH)
- rozpouštědlo pro spektroskopická měření: metanol (MeOH)
- absorpční maximum v metanolu: lexmax = 484 nm (molární extinkční koeficient: 154000 cm-1M-1)
- fluorescenční emisní maximum v metanolu: lemmax = 501 nm
- pracovní koncentrace se udává 40-200 nmol/l
- prochází buněčnou membránou
- může být použita pro značení jak mitochondrií, tak endoplazmatického retikula (po uvolnění sondy z mitochondrií)
Obr. 3.19 Potenciálová sonda DiOC6(3)
Materiál:
- DiOC6(3) jodid (Sigma)
- valinomycin (MW = 1111,36, Sigma)
- ouabain (MW = 584,2, Sigma)
- Triton X 100 (Merck)
- Ficoll-Paque Plus (Amersham Biosciences)
- centrifuga
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4) s 1% glukózou
- pufr B (120 mmol/l NaCl, 120 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4) s 1% glukózou
- skleněné zkumavky, mikropipety, …
Postup:
Izolace lymfocytů
z plné krve
1. krev odebrat do citrátového roztoku (nesrážlivá krev)
2. promíchat 10 ml krve a 10 ml pufru A
3. přidat 15 ml Ficoll-Paque Plus do průhledné centrifugační kyvety a opatrně navrstvit 20 ml zředěné krve
4.
centrifugovat při 400g po dobu 30 min při
5. odsát horní vrstvu (může být uchována pro pozdější použití)
6. přenést vrstvu s lymfocyty do centrifugační kyvety (cca 10 ml)
7. přidat alespoň 3 objemy pufru A, promíchat
8.
centrifugovat při 60-100g po 10 min při
9. odstranit supernatant a opakovat promytí
10. suspendovat lymfocyty v pufru A v koncentraci 5.107/ml
Měření změn
membránového potenciálu:
1. připravte zásobní roztok 10 mmol/l DiOC6(3) jodidu v DMSO a z něj pracovní roztok 0,1 mmol/l v etanolu
2. připravte tyto zásobní roztoky: 1 mmol/l valinomycin v etanolu, 2 mmol/l ouabain v pufru A, 10% Triton X 100 v pufru A
3. proveďte tato měření:
A) časová závislost fluorescence sondy po přidání k lymfocytům:
· připravte vzorky s výslednými koncentracemi 10 mmol/l DiOC6(3), 106 lymfocytů/ml
· změřte excitační a emisní spektra DiOC6(3) v samotném pufru A a v časech 1, 5, 10 a 30 minut po přidání lymfocytů
B) vliv depolarizace membrány na fluorescenci DiOC6(3):
· smíchejte jednotlivé složky podle Tabulky 3.7 ve výsledných koncentracích: 106 lymfocytů/ml, 0,2 mmol/l DiOC6(3), 1 mmol/l valinomycin, 100 mmol/l ouabain, 0,01% Triton X 100
· inkubujte ve tmě 30 min v 37 °C
·
promyjte dvakrát v pufru A nebo B
(centrifugací 10 min, 100 g,
·
s polarizátorem orientovaným svisle a
analyzátorem pootočeným o „magický“ úhel 54,74° změřte excitační a emisní
spektra nebo intenzitu fluorescence DiOC6(3) při buzení lexmax
= 484 nm a lemmax
= 519 nm; při
C) vliv Tritonu X 100 na fluorescenci DiOC6(3):
· změřte excitační a emisní spektra DiOC6(3) v pufru A bez lymfocytů v přítomnosti 0,01% a 0,1% Tritonu X 100
Tabulka 3.7
|
č. |
pufr A |
pufr B |
lymfocyty |
valinomycin |
ouabain |
Triton X 100 |
DiOC6(3) |
|
1 |
4890 |
0 |
100 |
0 |
0 |
0 |
10 |
|
2 |
4885 |
0 |
100 |
5 |
0 |
0 |
10 |
|
3 |
4640 |
0 |
100 |
0 |
250 |
0 |
10 |
|
4 |
4885 |
0 |
100 |
0 |
0 |
5 |
10 |
|
5 |
0 |
4890 |
100 |
0 |
0 |
0 |
10 |
|
6 |
0 |
4885 |
100 |
5 |
0 |
0 |
10 |
Výsledky:
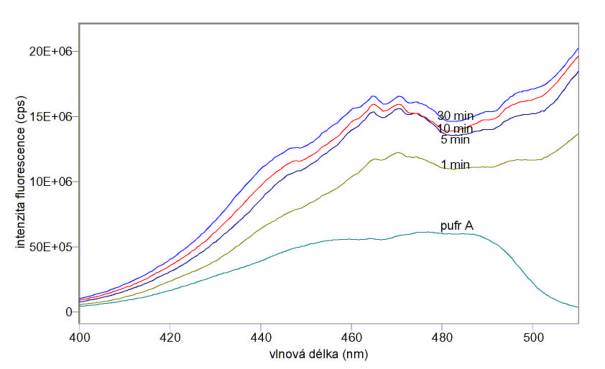
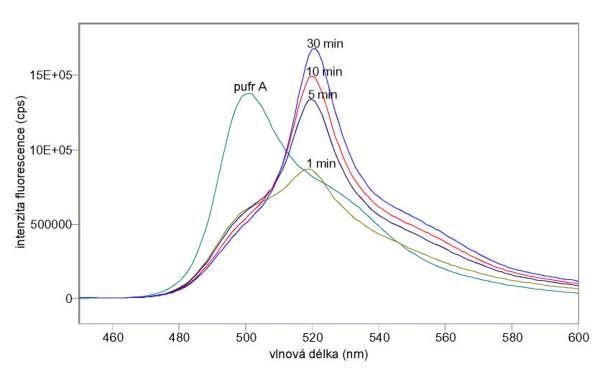
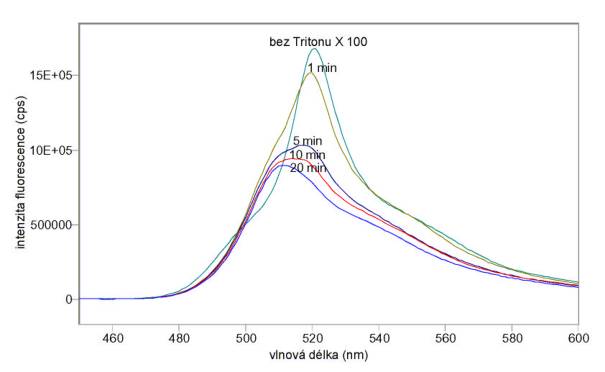
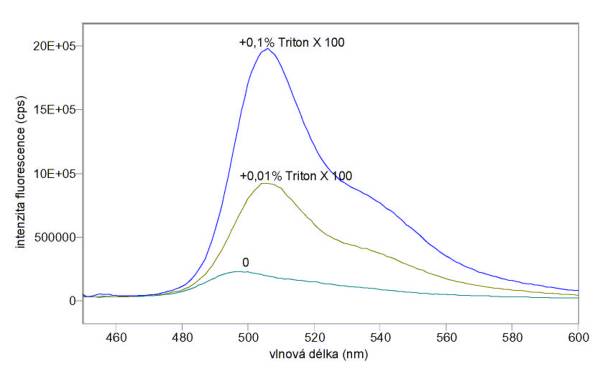
A) Na obr. 3.20 a 3.21 jsou excitační a emisní spektra sondy DiOC6(3) v koncentraci 10 mmol/l v samotném pufru A bez buněk a v různých časech po přidání lymfocytů v koncentraci 106/ml. Na obr. 3.22 jsou emisní spektra téhož vzorku v různých časech po přidání 0,01% Tritonu X 100. Při těchto měřeních nebyla odstraňována volná sonda.
B)
Na obr.
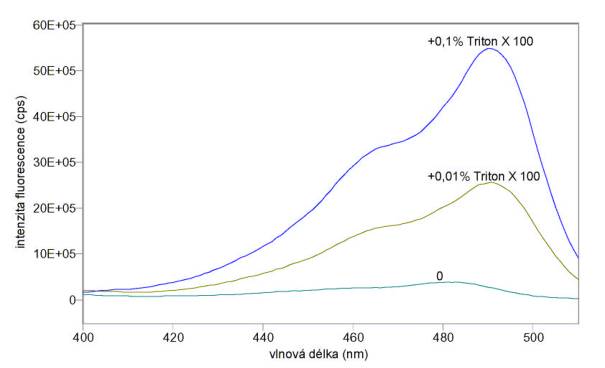
C) Na obr. 3.25 a 3.26 jsou excitační a emisní spektra sondy DiOC6(3) přidané k pufru A (bez buněk) v konečné koncentraci 200 nmol/l před a po přidání různých koncentrací Tritonu X 100. Výsledky ukazují, že samotný detergent velmi zvyšuje kvantový výtěžek fluorescence sondy DiOC6(3), což se musí brát v úvahu v experimentech, kdy Triton X 100 není dostatečně odmyt ze vzorku.
Obr. 3.20 Excitační spektra (lem = 520 nm) potenciálové sondy DiOC6(3) v pufru A před přidáním a v různých časech po přidání lymfocytů
Obr. 3.21 Emisní spektra (lex = 440 nm) potenciálové sondy DiOC6(3) v pufru A před přidáním a v různých časech po přidání lymfocytů
Obr. 3.22 Emisní spektra (lex = 440 nm) potenciálové sondy DiOC6(3) v pufru A s lymfocyty před přidáním a v různých časech po přidání 0,01% Tritonu X 100
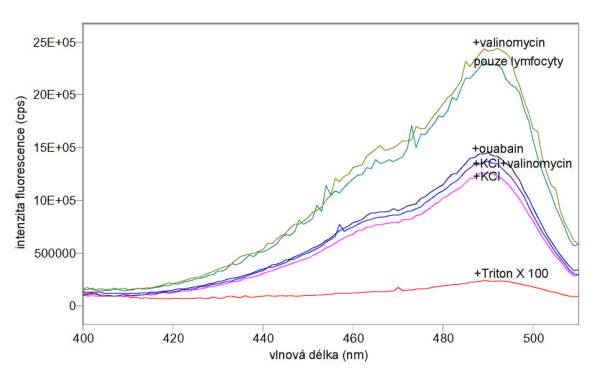
Obr. 3.23 Excitační spektra (lem = 520 nm) potenciálové sondy DiOC6(3) v lymfocytech inkubovaných: 1) v pufru A (pouze lymfocyty), 2) s 1 mmol/l valinomycinem v pufru A (+valinomycin), 3) se 100 mmol/l ouabainem v pufru A (+ouabain), 4) s 0,01% Tritonem X 100 v pufru A (+Triton X 100), 5) v pufru B (+KCl) a 6) v pufru B s 1 mmol/l valinomycinem (+KCl+valinomycin)
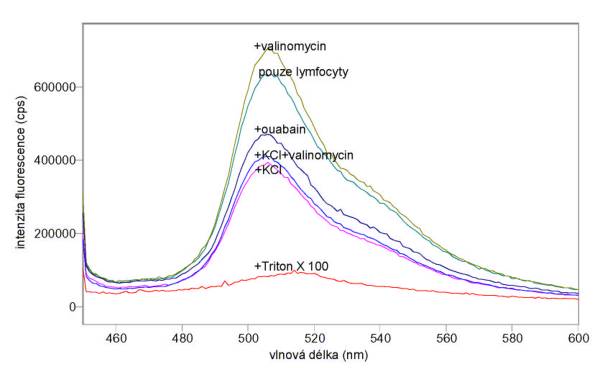
Obr. 3.24 Emisní spektra (lex = 440 nm) potenciálové sondy DiOC6(3) v lymfocytech inkubovaných: 1) v pufru A (pouze lymfocyty), 2) s 1 mmol/l valinomycinem v pufru A (+valinomycin), 3) se 100 mmol/l ouabainem v pufru A (+ouabain), 4) s 0,01% Tritonem X 100 v pufru A (+Triton X 100), 5) v pufru B (+KCl) a 6) v pufru B s 1 mmol/l valinomycinem (+KCl+valinomycin)
Obr. 3.25 Excitační spektra (lem = 520 nm) potenciálové sondy DiOC6(3) v pufru A při různých koncentracích Tritonu X 100
Obr. 3.26 Emisní spektra (lex = 440 nm) potenciálové sondy DiOC6(3) v pufru A při různých koncentracích Tritonu X 100
3.8. Změny pH
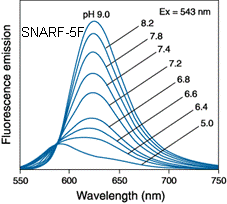
Sondy pro měření změn pH jsou popsány v kapitole 2.3.6..V tomto experimentu je ilustrována citlivost fluorescenčních sond pro měření pH. s použitím sondy SNARF-5F.
SNARF-5F patří do skupiny seminaftorhodafluorů, což je relativně nová skupina fluorescenčních indikátorů pH vyznačující se výraznými posuvy jak v absorpčním, tak v emisním spektru. Jedná se o sondy vhodné pro tzv. poměrná měření, kdy z poměru intenzit fluorescence měřené při dvou různých vlnových délkách emise lze zjistit pH okolního prostředí. Fluorescenční je jak kyselá, tak zásaditá forma těchto sond. Pro lepší prostupnost dovnitř živých buněk lze použít jejich acetoxymetylové (AM) nebo acetátové estery.
Vlastnosti sondy
SNARF-5F:
- MW = 471,44
- nízká rozpustnost ve vodě
- zásobní roztok: pH>6
- patří do skupiny seminaftorhodafluorů
- absorpční maximum v kyselém/neutrálním roztoku: lexmax = 555 nm (molární extinkční koeficient: 27000 cm-1M-1)
- fluorescenční emisní maximum v kyselém/neutrálním roztoku: lemmax = 590 nm
- absorpční maximum v zásaditém roztoku: lexmax = 579 nm (molární extinkční koeficient: 49000 cm-1M-1)
- fluorescenční emisní maximum v zásaditém roztoku: lemmax = 630 nm
- pKa = 7,2
Obr 3.27 Fluorescenční spektra sondy SNARF-5F při různých pH
(Podle: Handbook of
Fluorescent Probes and Research Products.)
Materiál:
- SNARF®-5F 5-(a 6)-karboxylová kyselina (Molecular Probes)
- skleněné zkumavky, mikropipety, …
- pufry s upraveným pH 5 až 9
Postup:
1. připravte zásobní roztok 2 mmol/l SNARF-5F v DMSO (dimetylsulfoxid)
2. do 5 ml pufru (pH = 5 – 9) přidejte (Hamiltonovou stříkačkou) 10 ml sondy SNARF-5F v konečné koncentraci 4 mmol/l
3. změřte excitační fluorescenční spektra vzorků při lem = 630 nm
4. změřte emisní fluorescenční spektra vzorků při lex = 540 nm
Výsledky:
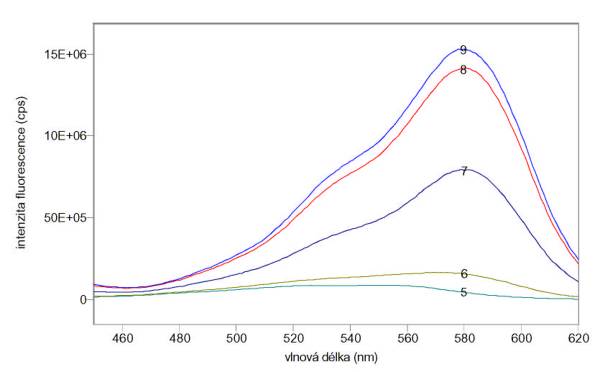
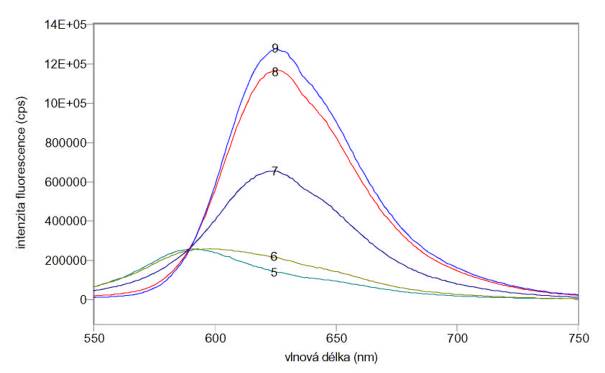
Změřená excitační a emisní spektra sondy SNARF-5F při různých pH jsou uvedena na obr. 3.28 a 3.29. Měření byla provedena na spektrofluorimetru FluoroMax-3.
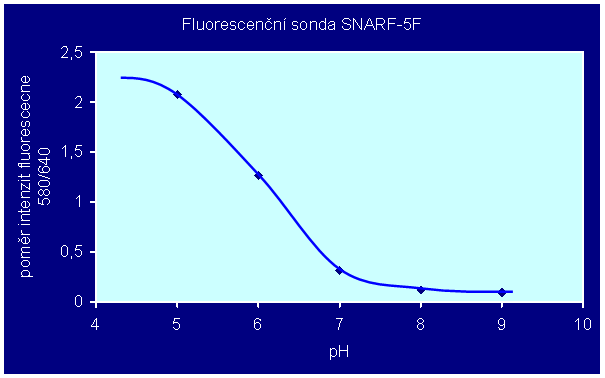
Sondy typu SNARF lze použít pro měření pH změřením poměru intenzit fluorescence při emisních vlnových délkách 580 a 640 nm; závislost poměru emisních intenzit na pH není lineární, jak je vidět z obr. 3.30. Kalibraci fluorescenční odezvy SNARF-5F lze provést pomocí pufrů s různými pH, v nichž je měřen poměr intenzit fluorescence při dvou různých vlnových délkách (l1=580 nm, l2=640 nm):
(3.1) pH = pKa – log[((R-RB)/(RA-R))*IB(l2)/IA(l2)]
kde je R poměr intenzit fluorescence při dvou různých vlnových délkách (I(l1)/I(l2)), indexy A a B označují limitní hodnoty v kyselé a zásadité oblasti titrační křivky; pro SNARF-5F je pKa=7,2. Korekce na fluorescenci pozadí musí provedena před výpočtem poměrů R.
Obr. 3.28 Excitační fluorescenční spektra 4 mmol/l SNARF-5F (lem = 630 nm) při různých hodnotách pH roztoku.
Obr. 3.29 Emisní fluorescenční spektra 4 mmol/l SNARF-5F (lex = 540 nm) při různých hodnotách pH roztoku.
Obr. 3.30 Závislost poměru intenzit fluorescence sondy SNARF-5F měřených při 580 a 640 nm na pH
3.9. Propustnost membrán
Karboxyfluorescein nebo calcein jsou zadržovány uvnitř neporušených buněk nebo lipidových vesiklů (viz též kap. 2.3.1). Fluorescence těchto látek je přitom >95% zhášena při koncentracích nad 100 mmol/l. Propustnost lipidových dvojvrstev pro takové fluorofory lze studovat tímto způsobem: koncentrovaný roztok fluoroforu ve vodě se uzavře v liposomech, které se potom oddělí od neuzavřeného barviva gelovou filtrací nebo centrifugací. Uvolňování fluoroforu z liposomů je potom provázeno zvyšováním intenzity fluorescence. Lze sledovat jak spontánní uvolňování, tak uvolňování vyvolané jinou látkou. Kompletní lýzu lze uskutečnit přidáním detergentu, Tritonu X-100.
Vlastnosti sondy 5-(a
6)-karboxyfluorescein (5(6)-FAM):
- MW = 376,32
- střední rozpustnost ve vodě; zásobní roztok v dimetylformamidu, pH>6
- absorpční maximum v kyselém/neutrálním roztoku: lexmax = 475 nm (molární extinkční koeficient: 28000 cm-1M-1)
- fluorescenční emisní maximum v kyselém/neutrálním roztoku: lemmax = 517 nm
- absorpční maximum v zásaditém roztoku: lexmax = 492 nm (molární extinkční koeficient: 75000 cm-1M-1)
- fluorescenční emisní maximum v zásaditém roztoku: lemmax = 517 nm
- pKa = 6,4
Materiál:
- 5-(a 6)-karboxyfluorescein (Fluka)
- fosfatidylcholin (Sigma)
- Triton X-100
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4)
- skleněné zkumavky, mikropipety, …
Postup:
1. připravte roztok 100 mmol/l 5-(a 6)-karboxyfluoresceinu v pufru A s pH zvýšeným na 9
2. připravte 10% roztok Tritonu-X v pufru A
3. připravte velké mnohovrstevné liposomy z fosfatidylcholinu, kdy jako vodný roztok použijte 100 mmol/l karboxyfluorescein:
- přenést
2 mg fosfolipidu ve směsi chloroform/metanol (2:1) do skleněné nádobky a
rozpouštědlo odpařit proudem dusíku při teplotě kolem
- nádobku umístit po několik hodin ve vakuu, aby byly odstraněny zbytky rozpouštědla
- přidat 2 ml 100 mmol karboxyfluoresceinu a vzorek inkubovat v 50 °C po dobu 5 minut
- intenzivně třepat a nakonec krátce sonikovat (cca 5 s) v sonikátoru XL 2020 (Heat Systems - Ultrasonics) na stupni 4
- inkubovat vzorek v 50 °C po dobu 30 minut a poté jej krátce protřepat
4.
liposomy promyjte 2 krát pufrem A (centrifugací při
30000 g, 15 min,
5. liposomy nařeďte pufrem A na výslednou koncentraci fosfatidylcholinu 100 mmol/ml
6.
v časech 30, 60, 90 a 120 minut po naředění
liposomů změřte excitační a emisní spektra nebo intenzitu fluorescence karboxyfluoresceinu
při buzení lex
= 490 nm a lem
= 520 nm; při
7. přidejte Triton X-100 v konečné koncentraci 0,1% a znovu změřte fluorescenci karboxyfluoresceinu
Výsledky:
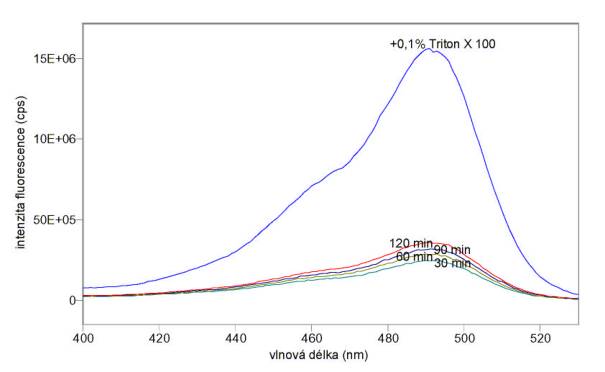
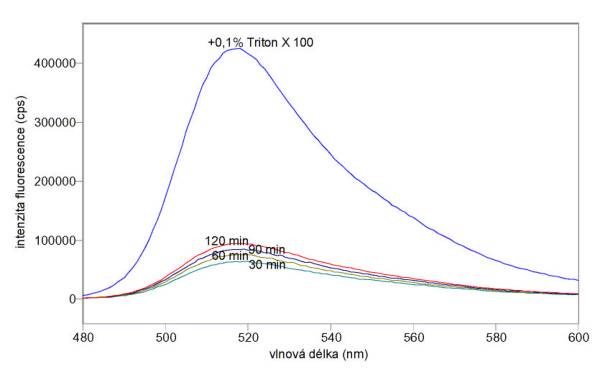
Na obr. 3.31 a 3.32 jsou excitační a emisní spektra 5-(a 6)-karboxyfluoresceinu uvolňovaného z mnohovrstevných fosfatidylcholinových liposomů obsahujících koncentrovaný fluorofor jednak spontánně (v časech 30, 60, 90 a 120 min po naředění liposomů pufrem A), jednak po destrukci liposomů 0,1% Tritonem X 100.
Obr 3.31 Excitační fluorescenční spektra (lem = 540 nm) 5-(a 6)-karboxyfluoresceinu uvolňovaného z mnohovrstevných liposomů spontánně v časech 30, 60, 90 a 120 minut po naředění liposomů a po úplném uvolnění působením 0,1% Tritonu X 100.
Obr 3.32 Emisní fluorescenční spektra (lex = 470 nm) 5-(a 6)-karboxyfluoresceinu uvolňovaného z mnohovrstevných liposomů spontánně v časech 30, 60, 90 a 120 minut po naředění liposomů a po úplném uvolnění působením 0,1% Tritonu X 100.
3.10. Speciální experimenty
3.10.1. Vazebné parametry receptorů
Membránové receptory pro neurotransmitery lze charakterizovat jejich afinitou Ka (nebo disociační konstantou Kd = 1/Ka) k neurotransmiteru či jinému ligandu a vazebnou kapacitou Bmax. Ka charakterizuje sílu vazby ligandu ke specifickému vazebnému místu na receptoru a Bmax odpovídá denzitě receptorů v membráně. Disociační konstanty neurotransmiterových receptorů leží obvykle v nanomolární oblasti a pro jejich měření tedy potřebujeme detekovat subnanomolární koncentrace vázajících se látek. Vazebné parametry receptorů se proto běžně určují pomocí ligandů, které k nim mají specifickou vazbu a které jsou značeny radionuklidy (obvykle agonisté nebo antagonisté receptorů). Metoda vazby radioligandu je velmi dobře rozpracována a popsána (např. v monografii [Neurotransmitter Receptor Binding, Yamamura, H.I., Enna, S.J. and Kuhar, M.J., eds., 2nd ed., Raven Press, New York, 1985]. Citlivost fluorescenční spektroskopie však umožňuje tato měření provádět i pomocí fluorescenčních ligandů (viz kap. 2.3.2), neboť za optimálních podmínek lze měřit i femtomolární koncentrace fluoroforů. Problém je dosud v nedostatečném počtu fluorescenčně značených ligandů pro neurotransmiterové receptory a větší pracnosti experimentu.
V tomto experimentu je popsán postup měření vazebných parametrů a1-adrenergních receptorů v lymfocytech pomocí jejich antagonisty - prazosinu fluorescenčně značeného látkou BODIPY FL. Postup je analogický postupu u metody vazby radioligandu, tj. měří se koncentrační závislost vazby ligandu k buněčným membránám. Pro oddělení volného a navázaného ligandu byla použita technika rychlé filtrace, která umožňuje velmi rychlé a snadné oddělení volného a vázaného ligandu za předpokladu, že použitý filtr je schopen zachytit částice, k nimž se ligand váže a současně je jeho propustnost dostatečná pro rychlý průchod promývací tekutiny. Pro záchyt lymfocytů jsme použili skleněné mikrovláknité filtry GF/C (Whatman), které byly impregnovány 0,1% polyetyleniminem pro snížení vazby volného ligandu na filtr. Vzorky byly naředěny 3 ml pufru A, filtrovány a filtry se zachycenými lymfocyty byly dvakrát promyty ledovým pufrem A (2 x 3 ml). Celková doba filtrace byla <10 sec., což umožňuje spolehlivé měření vazebných procesů s disociační konstantou <10-8 mol/l. Filtry byly umístěny do zkumavek a zhomogenizovány v 5 ml metanolu. Po centrifugaci byly 4 ml metanolu měřeny na spektrofluorimetru (lex = 490 nm, lem = 511 nm).
Nespecifická vazba byla určena jako vazba BODIPY FL prazosinu v přítomnosti nadbytku (10 mmol/l) neznačeného prazosinu. Tato vazba byla odečtena od celkové vazby a data byla vyhodnocena. Vyhodnocení dat je rovněž analogické jako při použití metody vazby radioligandu. Parametry vazebných izoterem Bmax a Kd byly spočteny metodami nelineární regresní analýzy s použitím programu AccuFit Saturation Two-Site (Beckman). Bylo použito označení běžné pro ligand-receptorové studie:
(3.2) B = Bmax . F / (Kd + F)
kde je B - množství prazosinu navázaného k 106 lymfocytů, F - koncentrace volného prazosinu, Bmax - maximální počet vazebných míst (vazebná kapacita) a Kd - rovnovážná disociační konstanta.
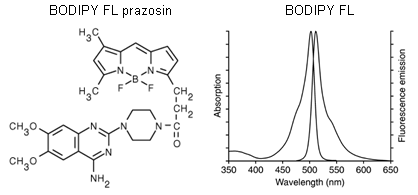
Vlastnosti BODIPY FL prazosinu:
- MW = 563,41
- rozpouštědlo pro zásobní roztok: dimetylsulfoxid (DMSO) nebo etanol (EtOH)
- rozpouštědlo pro spektroskopická měření: metanol (MeOH)
- absorpční maximum v metanolu: lexmax = 504 nm (molární extinkční koeficient: 77000 cm-1M-1)
- fluorescenční emisní maximum v metanolu: lemmax = 511 nm
- rozpustnost ve vodě je nízká
- antagonista a1-adrenoceptorů
Obr. 3.33 BODIPY FL prazosin a absorpční e emisní fluorescenční spektrum BODIPY FL.
(Podle: Handbook of
Fluorescent Probes and Research Products.)
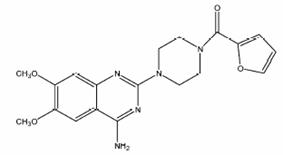
Obr. 3.34 Vzorec prazosinu
Materiál:
- BODIPY FL prazosin (Molecular Probes)
- prazosin (MW = 419,9, Sigma; rozpustný ve vodě do 0,5 mg/ml, v metanolu do 6 mg/ml)
- skleněné mikrovláknité filtry Whatman GF/C
- filtrační zařízení
- Ficoll-Paque Plus (Amersham Biosciences)
- metanol
- skleněné zkumavky, mikropipety, …
- centrifuga
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4)
Postup:
Izolace lymfocytů
z plné krve
- krev odebrat do citrátového roztoku (nesrážlivá krev)
- promíchat 2 ml krve a 2 ml pufru A
- přidat 3 ml Ficoll-Paque Plus do zkumavky a opatrně navrstvit 4 ml zředěné krve
- centrifugovat při 400g po
dobu 30 min při
- odsát horní vrstvu (může být uchována pro pozdější použití)
- přenést vrstvu s lymfocyty do centrifugační kyvety (cca 2 ml)
- přidat alespoň 3 objemy pufru A, promíchat
- centrifugovat při 60-100g
po 10 min při
- odstranit supernatant a opakovat promytí
- suspendovat lymfocyty v pufru A v koncentraci 5.107/ml
Vazba prazosinu na lymfocyty
1.
veškerá práce se dělá v ledové lázni (cca
2. připravit roztoky v pufru A:
- 50 mmol/l prazosin
- 100 nmol/l BODIPY FL prazosin
- 5 nmol/l BODIPY FL prazosin
3. do skleněných zkumavek pipetovat vzorky dle Tabulky 3.8; vazba se startuje přidáním BODIPY FL prazosinu
4. standardy: stejná množství BODIPY FL prazosinu jako byla přidána ke vzorkům pipetovat rovněž do prázdných vialek
5.
nechat inkubovat 30 min ve
6. filtry ještě dvakrát promýt 3 ml pufru A a potom je umístit do skleněných vialek
7. k filtrům i ke standardům přidat 5 ml metanolu; filtry homogenizovat (turaxovat), nechat přes noc ve tmě, centrifugovat a odebrat supernatant pro měření
8. měřit fluorescenci BODIPY FL prazosinu v metanolu (lex = 490 nm, lem = 511 nm)
9. analýza dat: spočítat parametry Kd a Bmax pomocí nelineární regrese (analogicky, jako se dělá v metodě vazby radioligandu)
Tabulka 3.8
|
č. vzorku |
pufr A |
lymfocyty (5.107/ml) |
prazosin (50 mmol/l) |
BODIPY FL prazosin (5 nmol/l) |
BODIPY FL prazosin (100 nmol/l) |
výsledná koncentrace BODIPY FL prazosinu (nmol/l) |
|
celková vazba: |
||||||
|
1 |
185 |
50 |
|
15 |
|
0,3 |
|
2 |
170 |
50 |
|
30 |
|
0,6 |
|
3 |
150 |
50 |
|
50 |
|
1 |
|
4 |
100 |
50 |
|
100 |
|
2 |
|
5 |
50 |
50 |
|
150 |
|
3 |
|
6 |
190 |
50 |
|
|
10 |
4 |
|
7 |
185 |
50 |
|
|
15 |
6 |
|
8 |
175 |
50 |
|
|
25 |
10 |
|
9 |
150 |
50 |
|
|
50 |
20 |
|
10 |
100 |
50 |
|
|
100 |
40 |
|
nespecifická vazba: |
||||||
|
11 |
135 |
50 |
50 |
15 |
|
0,3 |
|
12 |
120 |
50 |
50 |
30 |
|
0,6 |
|
13 |
100 |
50 |
50 |
50 |
|
1 |
|
14 |
50 |
50 |
50 |
100 |
|
2 |
|
15 |
0 |
50 |
50 |
150 |
|
3 |
|
16 |
140 |
50 |
50 |
|
10 |
4 |
|
17 |
135 |
50 |
50 |
|
15 |
6 |
|
18 |
125 |
50 |
50 |
|
25 |
10 |
|
19 |
100 |
50 |
50 |
|
50 |
20 |
|
20 |
50 |
50 |
50 |
|
100 |
40 |
|
standardy: |
||||||
|
21 |
|
|
|
15 |
|
|
|
22 |
|
|
|
30 |
|
|
|
23 |
|
|
|
50 |
|
|
|
24 |
|
|
|
100 |
|
|
|
25 |
|
|
|
150 |
|
|
|
26 |
|
|
|
|
10 |
|
|
27 |
|
|
|
|
15 |
|
|
28 |
|
|
|
|
25 |
|
|
29 |
|
|
|
|
50 |
|
|
30 |
|
|
|
|
100 |
|
Výsledky:
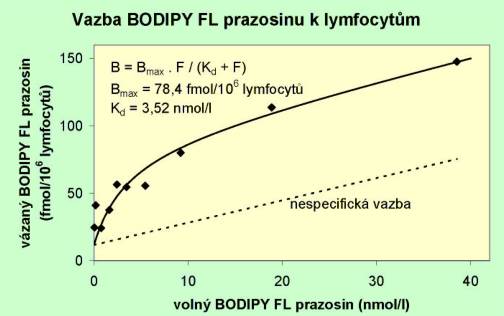
Na obr 3.35 je vynesena závislost BODIPY FL prazosinu vázaného na lymfocyty proti koncentraci volného (nenavázaného) BODIPY FL prazosinu. Tečkovaně je znázorněna nespecifická vazba, tj. vazba v přítomnosti nadbytku neznačeného prazosinu. Plná čára odpovídá rovnici B = 78,4 . F / (3,52 + F), kde B označuje vázaný a F volný BODIPY FL prazosin. V tomto experimentu tedy vyšla hodnota disociační konstanty pro vazbu prazosinu k lymfocytům Kd = 3,52 nmol/l a vazebná kapacita Bmax = 78,4 fmol/106 lymfocytů.
Obr. 3.35 Závislost BODIPY FL prazosinu vázaného k lymfocytům na koncentraci volného (mimobuněčného) BODIPY FL prazosinu.
3.10.2. Vliv psychotropních látek
Pro in vitro studium interakcí léčiv s lipidovou částí buněčných membrán jsou používány izolované buňky, membrány a modelové systémy. Rozdělení molekul mezi vodnou fázi a buněčné membrány je modelováno třemi základními systémy: 1. jednoduché rozdělení v systému voda/organické rozpouštědlo; 2. chromatografické rozdělovací systémy; 3. liposomy. Liposomy jsou široce používaným modelem lipidové části buněčných membrán a umožňují měřit nejen rozdělovací koeficienty, ale i vazebné parametry. Liposomy jsou váčky (vezikuly) uzavírající vodný roztok membránou tvořenou především fosfolipidy. Tvoří se spontánně, když jsou fosfolipidy dispergovány ve vodném prostředí. V závislosti na způsobu přípravy mohou vzniknout liposomy 1. mnohovrstevné (multilamellar vesicles, MLV), 2. malé jednovrstevné (small unilamellar vesicles, SUV) nebo 3. velké jednovrstevné (large unilamellar vesicles, LUV). V našich experimentech jsme použili MLV, které jsou tvořeny vezikuly o velikosti 10-1 až 100 mm (přičemž každý váček je tvořen mnoha koncentrickými vrstvičkami lipidových dvojvrstev) a LUV o průměru cca 100 nm. V mnoha experimentech bylo prokázáno, že MLV a LUV jsou dobrým modelem lipidové části buněčných membrán.
Metodou fluorescenčních sond s použitím membránové sondy 1,6-difenyl-1,3,5-hexatrien (DPH) byl měřen vliv různých koncentrací (10 nmol/l – 100 mmol/l) psychotropních látek na mikroviskozitu lipidových membrán připravených z fosfatidylcholinu (PC-LUV, PC-MLV):
- imipramin (IMI)
- amitriptylin (AMI)
- dothiepin (PRO)
- citalopram (CIT)
- chlorpromazin (CPR)
- klozapin(KLO)
- anandamid (ANA)
Materiál:
- 1,6-difenyl-1,3,5-hexatrien (DPH, Sigma)
- fosfatidylcholin (PC, Sigma)
- psychotropní látky (různý původ)
- zařízení LiposoFast-Basic
- skleněné zkumavky
- mikropipety
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4)
Postup:
Postup přípravy liposomů a měření se sondou DPH byl analogický jako je popsáno v kapitole 3.3.
Výsledky:
- výrazné snížení anizotropie fluorescence (zvýšení fluidity) bylo pozorováno při vyšších koncentracích anandamidu
- bez výrazné změny bylo přidání dothiepinu, amitriptylinu a citalopramu
- zvýšená anizotropie fluorescence DPH v membránách byla zjištěna vlivem imipraminu, chlorpromazinu a klozapinu
- s PC-MLV byly pozorovány podobné, pouze méně výraznější, změny rDPH jako s PC-LUV
Tabulka 3.9 Relativní změny anizotropie fluorescence sondy DPH v PC-LUV vyvolané různými koncentracemi psychotropních látek (imipramin (IMI), amitriptylin (AMI), dothiepin (PRO), citalopram (CIT), chlorpromazin (CPR), klozapin(KLO), anandamid (ANA))
|
Procento změny anizotropie fluorescence DPH v PC-LUV při
různých koncentracích psychotropních látek |
|||||||
|
konc. (mmol/l) |
IMI |
AMI |
PRO |
CIT |
CPR |
KLO |
ANA |
|
0,01 |
-0,09% |
-0,09% |
-1,05% |
-1,00% |
-1,25% |
-1,20% |
0,19% |
|
0,1 |
-0,03% |
-0,34% |
0,70% |
-3,70% |
0,27% |
1,19% |
-2,27% |
|
1 |
-0,32% |
0,34% |
-0,45% |
-2,46% |
2,16% |
2,57% |
-2,62% |
|
10 |
0,94% |
1,00% |
-2,23% |
-3,95% |
9,13% |
7,01% |
-8,41% |
|
100 |
6,86% |
-1,91% |
-1,22% |
-2,64% |
50,50% |
57,69% |
-29,21% |
3.10.3. Vliv podávání antidepresiv
Byly měřeny změny v lipidovém složení plazmatických membrán izolovaných z mozku a změny v jejich mikroviskozitě po dlouhodobém podávání antidepresiv. Laboratorním potkanům byla podávána antidepresiva s různými primárními biochemickými účinky (desipramin, maprotilin, citalopram, moklobemid a lithium) po dobu 4 týdnů. Byly izolovány plazmatické membrány z mozku. Chromatografickou analýzou lipidů z mozkových membrán bylo zjištěno významné snížení relativního zastoupení jednotlivých fosfolipidů vzhledem k celkovým lipidům (fosfolipidy + cholesterol) především po podávání maprotilinu, citalopramu a lithia. Toto snížení bylo způsobeno zvýšením koncentrací cholesterolu. Zvýšení obsahu cholesterolu v mozkových membránách může vést ke změnám v serotonergní neurotransmisi. Naše výsledky podporují hypotézu, že dlouhodobé podávání antidepresiv může způsobit změny v lipidovém složení buněčných membrán, které se mohou transformovat do procesů přímo ovlivňujících serotonergní neurotransmisi v mozku.
Biofyzikální vlastnosti synaptických membrán (strukturu, uspořádání, mikroviskozitu) určují do značné míry esenciální nenasycené mastné kyseliny, cholesterol a glykolipidy. Poměrně malé změny ve struktuře membrán způsobené cholesterolem a zastoupením w-3, w-6 a dalších nenasycených mastných kyselin potom mohou modulovat nejen metabolismus biogenních aminů, jejich vazbu nebo zpětné vychytávání, ale i transdukci signálu, např. vlivem na aktivitu adenylátcykláz a jiných membránových enzymů. V tzv. biofyzikální hypotéze afektivních poruch se uvažuje především o úloze dokosahexaenové kyseliny (22:6w-3) a arachidonové kyseliny (20:4w-6) ve funkci neuronálních membrán. Byla formulována také hypotéza o vztahu mezi sérovými lipidy, depresí a aterosklerosou. Podle hypotézy o optimální fluiditě membrán závisí maximální receptorová odezva nejen na hustotě receptorových vazebných míst, ale i na jejich přístupnosti pro agonisty rozpuštěné ve vodné fázi a na difúzně kontrolované rychlosti, s níž může docházet ke vzájemným interakcím mezi jednotlivými membránovými složkami podílejícími se na přenosu signálu. Jak dostupnost, tak pohyblivost membránových proteinů přitom závisí na mikroviskozitě lipidové dvojvrstvy. Hypotéza o měnitelné afinitě receptorů vychází z pozorování, že k výrazným změnám v afinitě receptorů vedou rozdíly nejen v chemické struktuře receptoru, ale i v lokálním membránovém mikrookolí a v různých nitrobuněčných procesech. Změny v lipidovém složení plazmatických membrán tedy mohou ovlivňovat přenos signálu a podílet se na terapeutických účincích antidepresiv.
Změna lipidového složení membrán ovlivňuje také aktivitu membránového serotoninového přenašeče (SERT). Tomuto přenašeči je v serotoninových hypotézách afektivních poruch věnována značná pozornost, neboť primárním biochemickým účinkem dnes nejčastěji podávaných antidepresiv, selektivních inhibitorů reuptake serotoninu (SSRI), je inhibice jeho funkce. Bylo popsáno snížení aktivity, denzity a dostupnosti SERT při depresi, nicméně snížení počtu uptake míst pro 5-HT nelze považovat za prokázané. Obvykle jsou měřeny parametry kinetiky transportu 5-HT do trombocytů, o nichž se předpokládá, že odrážejí změny funkce serotonergního systému v mozku. Významným modulátorem funkce SERT je především membránový cholesterol, jehož nižší koncentrace vedou ke snížení aktivity SERT. Propojení změn vlastností buněčných membrán způsobených různým obsahem cholesterolu s aktivitou serotonergního systému by mohlo být mechanismem podílejícím se na zvýšení rizika vzniku deprese spojené se suicidálním chováním při nízkých hladinách sérového cholesterolu. Rovněž snížená spotřeba w-3 nenasycených mastných kyselin koreluje se zvýšením rizika vzniku deprese.
Metody:
Desipramin, maprotilin, citalopram, moklobemid a lithium byly podávány žaludeční sondou v dávkách 10, 10, 5, 25 a 50 mg/kg.den po dobu 4 týdnů. Poté byla zvířata usmrcena a z mozku byly izolovány plazmatické membrány metodou dle Scottové a kol. (1993) s naší modifikací; obohacení vzorků plazmatickými membránami bylo kontrolováno stanovováním aktivity ekto-5’-nukleotidázy (Mitchell a Hawthorne 1965). Relativní změny fluidity membrán byly měřeny metodou fluorescenčních sond. Lipidová část membrán byla izolována modifikovanou Folchovou metodou (Folch et al. 1957; Koul a Prasad 1996). Lipidy byly analyzovány tenkovrstevnou chromatografií s plamenoionizační detekcí (TLC-FID) na zařízení Iatroscan TH10 nebo dvourozměrnou tenkovrstevnou chromatografií na deskách s následným stanovením fosforu a cholesterolu.
Chemikálie a roztoky:
- pufr
STM s inhibitory proteáz pro izolaci plazmatických membrán: 0,32 M
sacharóza, 10 mmol/l TRIS-HCl, 1 mmol/l MgCl2, 0,5 mmol/l PMSF,
0,5 mmol/l benzamidin HCl, 1 mmol/l EDTA, 2 mmol/l DTT (dithiothreitol),
pH 7,4. Pufr byl připraven den předem, po úpravě pH filtrován přes 0,45 µM
filtr a uložen ve
- pufr A (120 mmol/l NaCl, 10 mmol/l KCl, 30 mmol/l Tris HCl, pH 7,4).
Metoda
fluorescenčních sond
Pro měření relativních změn fluidity plazmatických membrán byla použita metoda fluorescenčních sond s hydrofobní membránovou sondou 1,6-difenyl-1,3,5-hexatrien (DPH, Sigma) a se sondou 1-(4-trimetylamonium-fenyl)-6-fenyl-1,3,5-hexatrien (TMA-DPH, Molecular Probes). DPH byl rozpuštěn v acetonu v koncentraci 6 mmol/l a TMA-DPH v metanolu v koncentraci 3 mmol/l a tyto zásobní roztoky byly uloženy v mrazničce. Pro měření byly připraveny vždy čerstvé roztoky sond v pufru A v koncentraci 6 µM (injektováním pomocí Hamiltonovy stříkačky 10 ml za intenzivního míchání). Tyto roztoky byl smíchány s pufrem A a suspenzí liposomů nebo membrán tak, aby výsledná koncentrace sondy byla 2 µM a koncentrace fosfolipidů cca 100 µmol/l. Po 60 minutách inkubace v 37°C byla měřena polarizovaná fluorescence (anizotropie fluorescence) při excitační vlnové délce 350 nm a emisní vlnové délce 450 nm na spektrofluorimetru FluoroMax-3 (Jobin Yvon, Horiba) s polarizačním příslušenstvím. Měření daného vzorku bylo opakováno alespoň 10x a hodnoty byly zprůměrňovány.
Polarizovaná fluorescence roztoků se měří ve směru kolmém ke směru
budícího paprsku, který je polarizován ve svislém směru. Je přitom nutno
provádět korekci na vliv emisního monochromátoru spektrofluorimetru na
polarizaci jím procházejícího záření. Anizotropie
fluorescence se určuje ze vztahu
(1.21) r
= (IIIV - G I^V)/((IIIV + 2 G I^V)
kde IIIV a I^V jsou složky světelné intenzity rovnoběžné nebo kolmé k
směru (vertikálnímu) polarizace budícího záření a G je korekční faktor, který
lze změřit při excitačním záření polarizovaném vodorovně jako poměr I^H/IIIH.
Izolace membránových
lipidů
Pro izolaci membránových lipidů byla použita modifikovaná metoda podle Folch a kol. (1957) a Koul a Prasad (1996). Stručný postup: vzorky byly turaxovány a po odebrání částí na stanovení koncentrace fosforu (Bartlett 1959, Wagner a kol. 1962) a proteinů (Lowryho et al. 1951) byly centrifugovány 10 min., 20000 g. Supernatant byl odstraněn a pelet roztřepán ve 20 objemech směsi dichlormetan:metanol (2:1, v/v) okyselené 0,25% HCl. V dusíkové atmosféře byly zkumavky jemně míchány v suché lázni při pokojové teplotě po dobu 15-30 min. Denaturované proteiny byly odstraněny filtrací přes gázu a filtrát (cca 5 ml) byl převeden do kalibrované zkumavky, přidány 0,2 objemu vody (cca 1 ml) a promícháno. Centrifugací 5 min. při 1000 g se oddělily fáze. Horní vrstva byla pečlivě odstraněna, čistá dolní fáze přenesena do předvážené očíslované lahvičky a odpařena pod dusíkem. Lahvičku se vzorkem byla zvážena a dána do mrazničky pod dusíkem. V celém postupu je nutno se vyhnout použití umělých hmot a minimalizovat přístup kyslíku (probublávat dusíkem).
Analýza membránových
lipidů
Pro analýzu membránových lipidů byla použita tenkovrstevná chromatografie, a to buď dvourozměrná chromatografie na skleněných deskách s následným stanovením fosforu a cholesterolu, nebo chromatografie na tyčinkách Chromarods 3 s plamenoionizační detekcí (TLC-FID) na zařízení Iatroscan TH10 (Iatron Laboratories).
Pro dvourozměrnou chromatografii byly použity tyto vyvíjecí
směsi: a) pro 1. směr – chloroform : metanol : NH4OH : H2O
v poměru 70 : 25 : 4 : 1; b) pro 2. směr – chloroform : metanol : aceton :
kyselina octová : H2O v poměru 70 : 12,5 : 17,5 : 10 : 4,5.
Chromatografické desky byly připraveny nanesením 0,5 mm vrstvy směsi 45 g
silikagelu H +
Pro chromatografii s plamenoionizační detekcí byly použity tyto vyvíjecí směsi: a) pro neutrální lipidy - hexan : éter : kys. mravenčí v poměru 45 : 15 : 0,3; b) pro fosfolipidy I - chloroform : metanol : izopropanol : voda : etylacetát v poměru 20 : 6 : 17 : 4 : 12; c) pro fosfolipidy II - chloroform : metanol : 7 N NH4OH v poměru 40 : 20 : 3. Po nanesení 1 µl vzorku byly vyvíjeny tyčinky nejprve v soustavě pro neutrální lipidy po dobu 2 x 20 minut a po vyvinutí spáleny na zařízení Iatroscan asi 0,75 cm od startu; byly tak získány píky pro cholesterol; poté byly tyčinky dále vyvíjeny v soustavě pro fosfolipidy I po dobu 2 x 60 minut a potom v soustavě pro fosfolipidy II po dobu 30 minut. Po vysušení byly tyčinky spáleny na zařízení Iatroscan a vyhodnoceny píky pro jednotlivé fosfolipidy.
Výsledky:
Vybrané výsledky jsou uvedeny v Tabulkách 3.10 a 3.11. Jejich podrobnější analýza je předmětem diskuze v připravené publikaci.
Tabulka
3.10 Relativní zastoupení molárních koncentrací jednotlivých fosfolipidů nebo
cholesterolu v plazmatických membránách izolovaných z mozku
laboratorních potkanů po 4-týdenním podávání různých antidepresiv (vztaženo
k celkové molární koncentraci fosfolipidů a cholesterolu).
|
skupina |
PC |
PE |
SM |
PI |
PS |
SUM |
CH |
|
KONTROLY |
0.208 |
0.042 |
0.075 |
0.044 |
0.043 |
0.412 |
0.369 |
|
DESIPRAMIN |
0.202 |
0.036 |
0.076 |
0.038 |
0.033 |
0.386 |
0.331 |
|
MAPROTILIN |
0.167 |
**0.019 |
0.065 |
0.036 |
*0.023 |
*0.311 |
0.373 |
|
CITALOPRAM |
0.181 |
*0.024 |
0.061 |
0.041 |
0.047 |
0.353 |
0.402 |
|
MOCLOBEMID |
0.186 |
*0.024 |
0.068 |
0.039 |
0.048 |
0.364 |
0.374 |
|
LiCO3 |
0.166 |
0.031 |
0.063 |
0.033 |
0.040 |
0.333 |
0.419 |
Průměry
byly spočteny z 15 hodnot pro kontrolní skupinu a z 6 hodnot pro
každou další skupinu. PC - fosfatidylcholin; PE - fosfatidyletanolamin; SM -
sfingomyelin; PI - fosfatidylinositol; PS - fosfatidylserin; SUM - suma molárních
koncentrací (PC+PE+SM+PI+PS); CH - cholesterol. Suma cholesterolu a celkových fosfolipidů
(CH+PL) byla použita jako 100%. Jsou označeny hodnoty statisticky významně
odlišné od kontrol (*p<0.05; **p<0.01; Mannův-Whitneyův U test)
Tabulka
3.11 Anizotropie fluorescence DPH a TMA-DPH v plazmatických membránách
izolovaných z mozků laboratorních potkanů po 4-týdenním podávání různých
antidepresiv.
|
skupina |
rDPH |
rTMA-DPH |
|
KONTROLY |
0.2074 |
0.2547 |
|
DESIPRAMIN |
*0.2043 |
0.2529 |
|
MAPROTILIN |
0.2055 |
0.2540 |
|
CITALOPRAM |
0.2090 |
0.2570 |
|
MOCLOBEMID |
0.2079 |
0.2560 |
|
LiCO3 |
0.2054 |
0.2534 |
Průměry
byly spočteny z 15 hodnot pro kontrolní skupinu a z 6 hodnot pro
každou další skupinu. DPH - 1,6-difenyl-1,3,5-hexatrien; TMA-DPH -
1-(4-trimetylamoniumfenyl)-6-fenyl-1,3,5-hexatrien. Je označena hodnota
statisticky významně odlišná od kontroly (*p<0.05; Mannův-Whitneyův U test)
Literatura:
Bartlett, G.R., 1959. Phosphorus assay in column
chromatography. J. Biol. Chem. 234, 466-468.
Folch J.L.M., Stanley, G.H.S.,
Koul, A., Prasad, R., 1996. Extraction
of membrane lipids. In: Manual
on membrane lipids, Prasad, R. (Ed.), Springer-Verlag, Berlin,
Heidelberg, pp. 37-51.
Lowry, O.H., Rosbrough, N.J., Farr, A.L., Randall, R.J., 1951.
Protein measurement with
the Folin phenol reagent. J. Biol. Chem. 193, 265-275.
Mitchell, R.H., Hawthorne, J.N., 1965. The site of diphosphoinositide synthesis in
rat liver. Biochem. Biophys. Res. Commun.
21, 333-338.
Scott, L., Schell, M.J., Hubbard, A.L., 1993. In: Graham, J.M., Higgins,
J.A. (Eds.), Methods in Molecular Biology, Vol. 19, Biomembrane Protocols: I. Isolation and Analysis. Humana Press Inc., Totowa, NJ.
Wagner, H., Lissau, A., Holzi, J., Horammer, L., 1962.
The incorporation of 32P into inositolphosphatides of the rat brain. J. Lipid
Res. 3, 177-180.